Tillbaka Klinik Kliniskt neurofysiologiskt stöd. El Paso, TX. Kiropraktor, dr Dr. Alexander Jimenez diskuterar klinisk neurofysiologi. Dr. Jimenez kommer att utforska den kliniska betydelsen och funktionella aktiviteterna av perifera nervfibrer, ryggmärgen, hjärnstammen och hjärnan i samband med viscerala och muskuloskeletala sjukdomar. Patienterna kommer att få en avancerad förståelse för smärtans anatomi, genetik, biokemi och fysiologi i relation till olika kliniska syndrom. Näringsbiokemi relaterad till nociception och smärta kommer att införlivas. Och implementeringen av denna information i terapiprogram kommer att betonas.
Vårt team är mycket stolta över att ge våra familjer och skadade patienter endast beprövade behandlingsprotokoll. Genom att lära ut fullständig holistisk välbefinnande som en livsstil förändrar vi också inte bara våra patienters liv utan också deras familjer. Vi gör detta för att vi ska kunna nå så många El Pasoans som behöver oss, oavsett prisproblem. För svar på alla frågor du kan ha, ring Dr Jimenez på 915-850-0900.
"Kliniska beslutsregler, ryggmärgsklassificering och förutsägelse av behandlingsresultat: En diskussion av de senaste rapporterna i rehabiliteringslitteraturen"
Abstrakt
Kliniska beslutsregler är en allt vanligare förekomst i den biomedicinska litteraturen och representerar en strategi för att förbättra det kliniska beslutsfattandet för att förbättra effektiviteten och effektiviteten i sjukvården. Inom ramen för rehabiliteringsforskning har kliniska beslutsregler främst varit inriktade på att klassificera patienter genom att förutsäga deras behandlingssvar på specifika terapier. Traditionellt föreslår rekommendationer för utveckling av kliniska beslutsregler en flerstegsprocess (härledning, validering, konsekvensanalys) med den definierade metoden. Forskningsinsatser som syftar till att utveckla en diagnosbaserad klinisk beslutsregel har avvikit från denna konvention. Nyligen publicerade publikationer inom denna forskningslinje har använt den modifierade terminologidiagnosbaserade kliniska beslutsguiden. Ändringar av terminologi och metodik kring kliniska beslutsregler kan göra det svårare för kliniker att känna igen nivån av evidens som är förknippad med en beslutsregel och förstå hur denna evidens bör implementeras för att informera patientvården. Vi ger en kort översikt över utvecklingen av kliniska beslutsregler i samband med rehabiliteringslitteraturen och två specifika artiklar som nyligen publicerats i Chiropractic and Manual Therapies.
Kliniska prediktionsregler
Sjukvården har genomgått ett viktigt paradigmskifte mot evidensbaserad praktik. Ett tillvägagångssätt tänkt att förbättra det kliniska beslutsfattandet genom att integrera bästa tillgängliga bevis med klinisk expertis och patienternas preferenser.
Ytterst är målet med evidensbaserad praxis att förbättra sjukvården. Men översättningen av vetenskapliga bevis till praktiken har visat sig vara en utmanande strävan.
Kliniska beslutsregler (CDR), även kända som kliniska prediktionsregler, är allt vanligare i rehabiliteringslitteraturen.
Dessa är verktyg utformade för att informera kliniskt beslutsfattande genom att identifiera potentiella prediktorer för diagnostiska testresultat, prognos eller terapeutiskt svar.
I rehabiliteringslitteraturen används CDR oftast för att förutsäga en patients svar på behandlingen. De har föreslagits för att identifiera kliniskt relevanta undergrupper av patienter som uppvisar annars heterogena störningar som ospecifik nacke eller låg ryggont, vilket är perspektivet som vi tänker fokusera på.
Kliniska prediktionsregler
Förmågan att klassificera eller undergruppera patienter med heterogena sjukdomar som ryggsmärta har lyfts fram som en forskningsprioritet och följaktligen i fokus för mycket forskning. Attraktionskraften hos sådana klassificeringsmetoder är deras potential för förbättrad behandlingseffektivitet och effektivitet genom att matcha patienter med optimala terapier. Tidigare har patientklassificering förlitat sig på implicita tillvägagångssätt grundade i tradition eller osystematiska observationer. Användningen av CDRs för att informera om klassificering är ett försök till ett mer evidensdrivet tillvägagångssätt, mindre beroende av ogrundade teorier.
CDR utvecklas i en flerstegsprocess som involverar studier av härledning, validering och analys av påverkan, där var och en har ett definierat syfte och metodologiska kriterier. Som med alla former av bevis som används för att fatta beslut om patienter, är uppmärksamhet på lämplig studiemetodik avgörande för att bedöma de potentiella fördelarna med implementering.
Fördelar med kliniska prediktionsregler
Den kan ta emot fler faktorer än den mänskliga hjärnan kan ta hänsyn till
CDR / CPR-modellen kommer alltid att ge samma resultat (matematisk ekvation)
Det kan vara mer exakt än klinisk bedömning.
Kliniska användningar av kliniska prediktionsregler
I slutändan ligger användbarheten av en CDR inte i dess noggrannhet utan i dess förmåga att förbättra kliniska resultat och förbättra vårdens effektivitet.[15] Även när en CDR uppvisar bred validering, säkerställer detta inte att det kommer att förändra det kliniska beslutsfattandet eller att de förändringar som det producerar kommer att resultera i bättre vård.
De förändringar det ger kommer att resultera i bättre vård. McGinn et al.[2] identifierade tre förklaringar till felet i en CDR i detta skede. För det första, om läkarens bedömning är lika korrekt som ett CDR-informerat beslut, finns det ingen fördel med dess användning. För det andra kan tillämpningen av en CDR innebära besvärliga beräkningar eller procedurer som avskräcker läkare från att använda CDR. För det tredje kanske det inte är möjligt att använda CDR i alla miljöer eller omständigheter. Dessutom skulle vi inkludera verkligheten att experimentella studier kan involvera patienter som inte är helt representativa för dem som ses i rutinvård och att detta kan begränsa det faktiska värdet av en CDR. För att till fullo förstå nyttan av en CDR och dess förmåga att förbättra sjukvården är det därför nödvändigt att genomföra en pragmatisk undersökning av dess genomförbarhet och effekt när den tillämpas i en miljö som återspeglar verklig praxis. Detta kan göras med olika studiedesigner som randomiserade studier, klusterrandomiserade studier eller andra metoder som att undersöka effekten av en CDR före och efter dess implementering.
Förekomst av klassificeringsmetoder för patienter med ländersjukdomar med användning av McKenzie syndrom, smärta mönster, manipulation och stabilisering kliniska prediktionsregler.
Syfte var (1) för att bestämma andelen patienter med lumbaldebarker som kunde klassificeras vid intag av McKenzie syndromer (McK) och smärtmönsterklassificering (PPC) med hjälp av mekaniska diagnos och terapi (MDT) bedömningsmetoder, manipulation och stabilisering klinisk förutsägelse regler (HLR) och (2) för varje Man CPR- eller Stab CPR-kategori, bestämmer klassificeringsprevalens med McK och PPC.
HLR är sofistikerade probabilistiska och prognostiska modeller där en grupp identifierade patientegenskaper och kliniska tecken och symtom är statistiskt associerade med meningsfull förutsägelse av patientens resultat.
Två separata HLR utvecklades av forskare för att identifiera patienter som skulle svara positivt på manipulation.33,34 Flynn et al. utvecklade den ursprungliga manipulations-HLR med hjälp av fem kriterier, dvs inga symtom under knäet, nyligen debuterade symtom (<16 dagar), lågt rädsla-undvikande frågeformulär36 för arbete (<19), hypomobilitet i ländryggen och inre höft. rotation ROM (>35 för minst en höft).33
Flynns HLR modifierades därefter av Fritz et al. till två kriterier, som inkluderade inga symtom under knäet och nyligen debut av symtom (<16 dagar), som ett pragmatiskt alternativ för att minska klinikernas börda för att identifiera patienter i primärvården som mest sannolikt reagerar på stötmanipulation.34 positivt
"Potentia.l fallgropar av kliniska prediktionsregler"
Vad är kliniska prediktionsregler?
En klinisk prediktionsregel (CPR) är en kombination av kliniska fynd som statistiskt har visat meningsfull förutsägbarhet vid bestämning av ett utvalt tillstånd eller prognos för en patient som har fått en specifik behandling 1,2. HLR skapas med hjälp av multivariata statistiska metoder, är utformade för att undersöka prediktionsförmågan hos utvalda grupperingar av kliniska variabler3,4, och är avsedda att hjälpa kliniker att fatta snabba beslut som normalt kan vara föremål för underliggande fördomar5. Reglerna är algoritmiska till sin natur och involverar kondenserad information som identifierar det minsta antalet statistiskt diagnostiska indikatorer för det måltillstånd6.
Kliniska prediktionsregler utvecklas i allmänhet med hjälp av en 3-stegsmetod14. För det första har HLR härlett oss prospektivt-
använda multivariata statistiska metoder för att undersöka prediktionsförmågan hos utvalda grupperingar av kliniska variabler3. Det andra steget innebär att validera HLR i en randomiserad kontrollerad studie för att minska risken att de prediktiva faktorer som utvecklades under härledningsfasen valdes ut av en slump14. Det tredje steget innebär att genomföra en konsekvensanalys för att fastställa hur HLR förbättrar vården, minskar kostnaderna och exakt definierar det uppsatta målet14.
Även om det finns liten debatt om att omsorgsfullt konstruerade HLR kan förbättra klinisk praxis, så vet jag att det inte finns några riktlinjer som anger metodiska krav för HLR för infusion i alla kliniska praktikmiljöer. Riktlinjer skapas för att förbättra strängheten i studiedesign och rapportering. I följande redaktion beskrivs potentiella metodiska fallgropar i HLR som kan väsentligt försvaga överföringsförmågan hos algoritmen. Inom rehabiliteringsområdet har de flesta HLR varit receptfria. Därför är mina kommentarer här reflekterande av prescriptiva HLR.
Metodiska fallgropar
HLR är utformade för att specificera en homogen uppsättning egenskaper från en heterogen population av prospektivt utvalda på varandra följande patienter5,15. Vanligtvis är den resulterande tillämpliga populationen en liten delmängd av ett större urval och kan bara representera en liten andel av läkarens faktiska dagliga fallbelastning. Inställningen och placeringen av det större urvalet bör vara generaliserbart15,16, och efterföljande validitetsstudier kräver bedömning av HLR i olika patientgrupper, i olika miljöer och med en typisk patientgrupp som ses av de flesta läkare16. Eftersom många HLR är utvecklade baserat på en mycket distinkt grupp som kanske eller kanske inte speglar en typisk patientpopulation, kan spektrumtransporterbarheten17 för många nuvarande HLR-algoritmer vara begränsad.
Kliniska prediktionsregler använder resultatmått för att bestämma effektiviteten av interventionen. Resultatmått måste ha en enda operativ definition5 och kräva tillräcklig lyhördhet för att verkligen fånga lämplig förändring i tillståndet14; Dessutom bör dessa åtgärder ha en välkonstruerad cut-off poäng16,18 och samlas in av en blindad administratör15. Valet av en lämplig ankarpoäng för mätning av faktisk förändring diskuteras för närvarande19-20. De flesta utfallsmått använder ett patientåterkallelsebaserat frågeformulär såsom en global rating of change score (GRoC), som är lämplig när den används på kort sikt men lider av recall bias när den används i långtidsanalyser19-21.
En potentiell nackdel för HLR är misslyckandet med att upprätthålla kvaliteten på de tester och mätningar som används som prediktorer i algoritmen. Därför bör perspektivtestet och måtten vara oberoende av varandra under modellering16; var och en bör utföras på ett meningsfullt, acceptabelt sätt4; Kliniker eller dataadministratörer bör vara blinda för patientens resultatmått och tillstånd22.
Källor
Potentiella fallgropar av kliniska förutsägelsesregler; Journal of Manual & Manipulative Therapy Volume 16 Number Two [69]
Jeffrey J Hebert och Julie M Fritz; Kliniska beslutsregler, ryggmärgsklassificering och förutsägelse av behandlingsresultat: En diskussion av de senaste rapporterna i rehabiliteringslitteraturen
Depression är ett av de vanligaste psykiska hälsoproblemen i USA. Aktuell forskning tyder på att depression beror på en kombination av genetiska, biologiska, ekologiska och psykologiska aspekter. Depression är en allvarlig psykiatrisk störning över hela världen med en betydande ekonomisk och psykologisk påfrestning på samhället. Lyckligtvis kan depression, även de svåraste fallen, behandlas. Ju tidigare behandlingen kan påbörjas, desto effektivare är den.
Som ett resultat finns det dock ett behov av robusta biomarkörer som hjälper till att förbättra diagnosen för att påskynda läkemedels- och/eller läkemedelsupptäcktsprocessen för varje patient med sjukdomen. Dessa är objektiva, perifera fysiologiska indikatorer där närvaro kan användas för att förutsäga sannolikheten för att depression ska debutera eller förekomma, stratifiera enligt svårighetsgrad eller symptomatologi, indikera förutsägelse och prognos eller övervaka svar på terapeutiska ingrepp. Syftet med följande artikel är att visa nya insikter, nuvarande utmaningar och framtidsutsikter när det gäller upptäckten av en mängd olika biomarkörer för depression och hur dessa kan hjälpa till att förbättra diagnos och behandling.
Biomarkörer för depression: Nya insikter, aktuella utmaningar och framtidsutsikter
Abstrakt
En uppsjö av forskning har involverat hundratals förmodade biomarkörer för depression, men har ännu inte helt klarlagt deras roller i depressiv sjukdom eller fastställt vad som är onormalt hos vilka patienter och hur biologisk information kan användas för att förbättra diagnos, behandling och prognos. Denna brist på framsteg beror delvis på depressionens natur och heterogenitet, i kombination med metodologisk heterogenitet inom forskningslitteraturen och det stora utbudet av biomarkörer med potential, vars uttryck ofta varierar beroende på många faktorer. Vi granskar den tillgängliga litteraturen, som indikerar att markörer involverade i inflammatoriska, neurotrofiska och metaboliska processer, såväl som neurotransmittor och neuroendokrina systemkomponenter, representerar mycket lovande kandidater. Dessa kan mätas genom genetiska och epigenetiska, transkriptomiska och proteomiska, metabolomiska och neuroavbildningsbedömningar. Användningen av nya tillvägagångssätt och systematiska forskningsprogram krävs nu för att avgöra om och vilka biomarkörer som kan användas för att förutsäga behandlingssvar, stratifiera patienter till specifika behandlingar och utveckla mål för nya interventioner. Vi drar slutsatsen att det finns mycket lovande för att minska bördan av depression genom att vidareutveckla och utöka dessa forskningsvägar.
Även om psykiatrin har en sjukdomsrelaterad börda som är större än någon enskild annan medicinsk diagnostisk kategori,1 är en skillnad i aktning fortfarande uppenbar mellan fysisk och psykisk hälsa inom många områden, inklusive forskningsfinansiering2 och publicering.3 Bland svårigheterna som psykisk hälsa står inför är bristen konsensus kring klassificering, diagnos och behandling som härrör från en ofullständig förståelse av de processer som ligger bakom dessa störningar. Detta är mycket uppenbart i humörstörningar, den kategori som utgör den enskilt största bördan för psykisk hälsa.3 Den vanligaste sinnesstämningen, major depressive disorder (MDD), är en komplex, heterogen sjukdom där upp till 60 % av patienterna kan uppleva någon grad av behandlingsresistens som förlänger och förvärrar episoder.4 För humörstörningar och inom det bredare fältet av psykisk hälsa skulle behandlingsresultaten sannolikt förbättras genom upptäckten av robusta, homogena subtyper inom (och över) diagnostiska kategorier, genom vilka behandlingar skulle kunna stratifieras. Som ett erkännande av detta pågår nu globala initiativ för att avgränsa funktionella subtyper, såsom forskningsdomänkriterierna.5 Det har hävdats att biologiska markörer är prioriterade kandidater för att subtypa psykiska störningar.6
Förbättra svar på behandlingar för depression
Trots ett omfattande utbud av behandlingsalternativ för allvarlig depression uppnår endast cirka en tredjedel av patienterna med MDD remission även när de får optimal antidepressiv behandling enligt konsensusriktlinjer och använder mätningsbaserad vård, och graden av behandlingssvar verkar minska med varje ny behandling .7 Dessutom är behandlingsresistent depression (TRD) associerad med ökad funktionsnedsättning, mortalitet, sjuklighet och återkommande eller kroniska episoder på lång sikt.8,9 Att uppnå förbättringar av behandlingssvaret i alla kliniska skeden skulle alltså ge bredare fördelar för övergripande resultat vid depression. Trots den betydande börda som kan hänföras till TRD har forskningen på detta område varit sparsam. Definitioner av TRD är inte standardiserade, trots tidigare försök:4 vissa kriterier kräver endast en behandlingsprövning som inte lyckas uppnå en 50 % minskning av symtompoängen (från ett validerat mått på depressionens svårighetsgrad), medan andra kräver att full remission inte uppnås eller utebliven respons på minst två adekvat utprövade antidepressiva medel av olika klasser inom en episod för att betraktas som TRD.4,10 Dessutom förbättras stadieindelningen och förutsägelsen av behandlingsresistens genom att lägga till de viktigaste kliniska egenskaperna för svårighetsgrad och kroniskhet till antalet misslyckade behandlingar .9,11 Ändå gör denna inkonsekvens i definitionen tolkningen av forskningslitteraturen om TRD till en ännu mer komplex uppgift.
För att förbättra responsen på behandlingar är det helt klart till hjälp att identifiera prediktiva riskfaktorer för utebliven respons. Vissa allmänna prediktorer för TRD har karakteriserats, inklusive avsaknad av full remission efter tidigare episoder, komorbid ångest, suicidalitet och tidig depression, såväl som personlighet (särskilt låg extraversion, lågt belöningsberoende och hög neuroticism) och genetiska faktorer.12 Dessa fynd bekräftas av recensioner som syntetiserar bevisen separat för farmakologisk13 och psykologisk14 behandling av depression. Antidepressiva medel och kognitiva beteendeterapier visar ungefär jämförbar effekt,15 men på grund av deras olika verkningsmekanismer kan de förväntas ha olika prediktorer för svar. Även om trauma under tidiga liv länge har förknippats med sämre kliniska resultat och minskade behandlingssvar,16 tyder tidiga indikationer på att personer med en historia av barndomstrauma kan svara bättre på psykologiska än farmakologiska terapier.17 Trots detta råder osäkerhet och liten personalisering eller stratifiering av behandlingen har nått klinisk praxis.18
Denna recension fokuserar på bevis som stöder användbarheten av biomarkörer som potentiellt användbara kliniska verktyg för att förbättra behandlingssvaret för depression.
Biomarkörer: System och källor
Biomarkörer tillhandahåller ett potentiellt mål för att identifiera prediktorer för svar på olika ingrepp.19 Bevisen hittills tyder på att markörer som återspeglar aktiviteten hos inflammatoriska, neurotransmittorer, neurotrofa, neuroendokrina och metabola system kan kunna förutsäga mentala och fysiska hälsoresultat hos för närvarande deprimerade individer , men det finns mycket inkonsekvens mellan fynden.20 I denna översikt fokuserar vi på dessa fem biologiska system.
För att få en fullständig förståelse för molekylära vägar och deras bidrag vid psykiatriska störningar anses det nu vara viktigt att bedöma flera biologiska nivåer, i vad som populärt kallas för en omics-metod.21 Figur 1 ger en skildring av de olika biologiska nivåer vid vilka vart och ett av de fem systemen kan bedömas, och de potentiella källorna till markörer på vilka dessa bedömningar kan göras. Observera dock att även om varje system kan inspekteras på varje omics-nivå, varierar de optimala mätkällorna klart på varje nivå. Till exempel ger neuroimaging en plattform för indirekt bedömning av hjärnans struktur eller funktion, medan proteinundersökningar i blod direkt bedömer markörer. Transcriptomics22 och metabolomics23 blir allt populärare och erbjuder bedömning av potentiellt enorma antal markörer, och Human Microbiome Project försöker nu identifiera alla mikroorganismer och deras genetiska sammansättning inom människor.24 Nya teknologier förbättrar vår förmåga att mäta dessa, inklusive genom ytterligare källor ; till exempel kan hormoner som kortisol nu analyseras i hår eller fingernaglar (som ger en kronisk indikation) eller svett (som ger en kontinuerlig mätning),25 såväl som i blod, cerebrospinalvätska, urin och saliv.
Med tanke på antalet förmodade källor, nivåer och system som är involverade i depression är det inte förvånande att omfattningen av biomarkörer med translationspotential är omfattande. Särskilt när interaktioner mellan markörer övervägs är det kanske osannolikt att undersökning av enskilda biomarkörer isolerat kommer att ge resultat som är fruktbara för att förbättra klinisk praxis. Schmidt et al26 föreslog användningen av biomarkörpaneler och därefter skisserade Brand et al27 ett utkast till panel baserat på tidigare kliniska och prekliniska bevis för MDD, som identifierade 16 "starka" biomarkörsmål, som var och en sällan är en enda markör. De omfattar minskad volym av grå substans (i hippocampus, prefrontal cortex och basala ganglier), förändringar i dygnscykeln, hyperkortisolism och andra representationer av hyperaktivering av hypotalamus-hypofys-binjureaxeln (HPA), dysfunktion av sköldkörteln, reducerad dopamin, noradrenalin eller 5-hydroxiindol. ökad glutamat, ökad superoxiddismutas och lipidperoxidation, försvagad cyklisk adenosin-3a,5a-monofosfat- och mitogenaktiverad proteinkinasvägaktivitet, ökade proinflammatoriska cytokiner, förändringar av tryptofan, kynurenin, insulin och specifika genetiska polymorfismer. Dessa markörer har inte kommit överens med konsensus och skulle kunna mätas på olika sätt; det är tydligt att fokuserat och systematiskt arbete måste ta itu med denna enorma uppgift för att bevisa deras kliniska fördelar.
Syftet med denna recension
Som en medvetet bred översikt, försöker den här artikeln fastställa de övergripande behoven av biomarkörforskning vid depression och i vilken utsträckning biomarkörer har verklig translationspotential för att förbättra responsen på behandlingar. Vi börjar med att diskutera de viktigaste och mest spännande resultaten inom detta område och riktar läsaren till mer specifika recensioner som rör relevanta markörer och jämförelser. Vi skisserar de aktuella utmaningarna i ljuset av bevisen, i kombination med behov av att minska bördan av depression. Slutligen ser vi framåt mot de viktiga forskningsvägarna för att möta nuvarande utmaningar och deras konsekvenser för klinisk praxis.
Senaste insikter
Sökandet efter kliniskt användbara biomarkörer för personer med depression har genererat omfattande undersökningar under det senaste halvseklet. De vanligaste behandlingarna kom från monoaminteorin om depression; därefter fick neuroendokrina hypoteser stor uppmärksamhet. På senare år har den mest produktiva forskningen kring den inflammatoriska hypotesen om depression. Ett stort antal relevanta översiktsartiklar har dock fokuserat på alla fem systemen; se tabell 1 och nedan för en samling av nya insikter över biomarkörsystem. Även om de mäts på många nivåer, har blodhärledda proteiner undersökts mest och ger en källa till biomarkörer som är bekväm, kostnadseffektiv och kan vara närmare translationspotential än andra källor; sålunda ges mer detaljer om biomarkörer som cirkulerar i blodet.
I en nyligen genomförd systematisk översikt undersökte Jani et al20 perifera blodbaserade biomarkörer för depression i samband med behandlingsresultat. Av endast 14 inkluderade studier (sökte fram till början av 2013) studerades 36 biomarkörer, varav 12 var signifikanta prediktorer för mentala eller fysiska responsindex i minst en undersökning. De som identifierats som potentiellt representerande riskfaktorer för utebliven respons inkluderade inflammatoriska proteiner: lågt interleukin (IL)-12p70, förhållandet mellan lymfocyter och monocyter; neuroendokrina markörer (dexametason icke-suppression av kortisol, högt cirkulerande kortisol, reducerat sköldkörtelstimulerande hormon); signalsubstansmarkörer (lågt serotonin och noradrenalin); metaboliska (låg högdensitetslipoproteinkolesterol) och neurotrofiska faktorer (reducerad S100 kalciumbindande protein B). Utöver detta har andra recensioner rapporterat om samband mellan ytterligare biomarkörer och behandlingsresultat.19,28�30 En kort beskrivning av förmodade markörer i varje system beskrivs i de efterföljande avsnitten och i Tabell 2.
Inflammatoriska fynd vid depression
Sedan Smiths framträdande artikel som beskriver makrofaghypotesen31 har denna etablerade litteratur funnit ökade nivåer av olika proinflammatoriska markörer hos deprimerade patienter, vilka har granskats flitigt.32�37 Tolv inflammatoriska proteiner har utvärderats i metaanalyser som jämförde deprimerade och friska kontrollpopulationer.38�43
IL-6 (P<0.001 i alla metaanalyser; 31 studier inkluderade) och CRP (P<0.001; 20 studier) uppträder ofta och tillförlitligt förhöjda vid depression.40 Förhöjd tumörnekrosfaktor alfa (TNF?) identifierades i tidiga studier (P<0.001),38 men betydande heterogenitet gjorde detta ofullständigt när man tar hänsyn till nyare undersökningar (31 studier).40 IL-1? är ännu mer osäkert förknippat med depression, med metaanalyser som tyder på högre nivåer av depression (P=0.03),41 höga nivåer endast i europeiska studier42 eller inga skillnader från kontroller.40 Trots detta föreslog en nyligen publicerad artikel särskilda translationella implikationer för IL- 1?,44 stöds av en extremt signifikant effekt av förhöjd IL-1? ribonukleinsyra som förutsäger ett dåligt svar på antidepressiva medel;45 andra fynd ovan avser cirkulerande blod-härledda cytokiner. Kemokinmonocyten kemoattraktant protein-1 har visat förhöjningar hos deprimerade deltagare i en metaanalys.39 Interleukinerna IL-2, IL-4, IL-8, IL-10 och interferon gamma var inte signifikant olika mellan deprimerade patienter och kontroller vid en metaanalytisk nivå, men har ändå visat potential när det gäller att förändras med behandling: IL-8 har rapporterats som förhöjt hos personer med svår depression prospektivt och tvärsnittsmässigt, 46 olika mönster av förändring i IL-10 och interferon gamma under behandling har förekommit mellan patienter som svarar tidigt mot personer som inte svarar,47 medan IL-4 och IL-2 har minskat i takt med symtomremission.48 I metaanalyser har små minskningar vid sidan av behandlingen påvisats för IL-6, IL-1?, IL- 10 och CRP.43,49,50 Dessutom TNF? kan bara minska med behandling hos responders, och ett sammansatt markörindex kan indikera ökad inflammation hos patienter som därefter inte svarar på behandlingen.43 Det är dock anmärkningsvärt att nästan all forskning som undersöker inflammatoriska proteiner och behandlingssvar använder farmakologiska behandlingsförsök. . Sålunda kan åtminstone vissa inflammatoriska förändringar under behandlingen troligen tillskrivas antidepressiva medel. De exakta inflammatoriska effekterna av olika antidepressiva medel har ännu inte fastställts, men bevis som använder CRP-nivåer tyder på att individer reagerar olika på specifika behandlingar baserat på baslinjeinflammation: Harley et al51 rapporterade förhöjd CRP för behandling som förutsäger ett dåligt svar på psykologisk terapi (kognitiv beteende eller interpersonell psykoterapi), men ett bra svar på nortriptylin eller fluoxetin; Uher et al52 replikerade detta fynd för nortriptylin och identifierade den motsatta effekten för escitalopram. Däremot fann Chang et al53 högre CRP hos patienter med tidiga svar på fluoxetin eller venlafaxin än hos icke-svarare. Dessutom har patienter med TRD och hög CRP svarat bättre på TNF? antagonist infliximab än de med nivåer inom normalområdet.54
Tillsammans tyder bevisen på att även när man kontrollerar för faktorer som kroppsmassaindex (BMI) och ålder, verkar inflammatoriska svar avvikande hos ungefär en tredjedel av patienterna med depression.55,56 Det inflammatoriska systemet är dock extremt komplext och det finns många biomarkörer som representerar olika aspekter av detta system. Nyligen har ytterligare nya cytokiner och kemokiner gett bevis på abnormiteter vid depression. Dessa inkluderar: makrofaghämmande protein 1a, IL-1a, IL-7, IL-12p70, IL-13, IL-15, eotaxin, granulocytmakrofagkolonistimulerande faktor,57 IL-5,58 IL-16,59 IL- 17,60 monocyt kemoattraherande protein-4,61 tymus och aktiveringsreglerad kemokin,62 eotaxin-3, TNFb,63 interferon gamma-inducerat protein 10,64 serumamyloid A,65 löslig intracellulär adhesionsmolekyl66 och löslig molekyl 1.67 kärlcell och löslig XNUMX kärlcell
Tillväxtfaktorfynd vid depression
I ljuset av den potentiella betydelsen av icke-neurotrofa tillväxtfaktorer (som de som är relaterade till angiogenes), hänvisar vi till neurogena biomarkörer under den bredare definitionen av tillväxtfaktorer.
Brain-derived neurotrophic factor (BDNF) är den mest studerade av dessa. Flera metaanalyser visar försvagningar av BDNF-proteinet i serum, som tycks öka tillsammans med antidepressiv behandling.68�71 Den senaste av dessa analyser tyder på att dessa BDNF-avvikelser är mer uttalade hos de mest allvarligt deprimerade patienterna, men att antidepressiva läkemedel verkar vara mer uttalade. öka nivåerna av detta protein även i frånvaro av klinisk remission.70 proBDNF har studerats mindre brett än den mogna formen av BDNF, men de två verkar skilja sig åt funktionellt (när det gäller deras effekter på tyrosinreceptorkinas B-receptorer) och nyligen bevis tyder på att även om mogen BDNF kan reduceras vid depression, kan proBDNF överproduceras.72 Nervtillväxtfaktor bedömd perifert har också rapporterats som lägre vid depression än hos kontroller i en metaanalys, men kanske inte förändras av antidepressiv behandling trots att den är mest försvagat hos patienter med svårare depression.73 Liknande fynd har rapporterats i en metaanalys för gliacellerlinjehärledd neurotrofisk faktor.74
Vaskulär endoteltillväxtfaktor (VEGF) har en roll för att främja angiogenes och neurogenes tillsammans med andra medlemmar av VEGF-familjen (t.ex. VEGF-C, VEGF-D) och har lovande för depression.75 Trots inkonsekventa bevis har två metaanalyser nyligen indikerade förhöjningar av VEGF i blod från deprimerade patienter jämfört med kontroller (över 16 studier; P<0.001).76,77 Låg VEGF har dock identifierats i TRD78 och högre nivåer har förutspått utebliven respons på antidepressiv behandling.79 Det är inte förstått varför nivåerna av VEGF-protein skulle vara förhöjda, men det kan delvis bero på proinflammatorisk aktivitet och/eller ökning av blodhjärnbarriärpermeabiliteten i deprimerade tillstånd som orsakar minskat uttryck i cerebrospinalvätskan.80 Sambandet mellan VEGF och behandlingssvar är oklart ; en nyligen genomförd studie fann inget samband mellan vare sig serum VEGF eller BDNF med respons eller svårighetsgrad av depression, trots minskningar vid sidan av antidepressiv behandling.81 Insulinliknande tillväxtfaktor-1 är en ytterligare faktor med neurogena funktioner som kan öka vid depression, vilket återspeglar en obalans i neurotrofiska processer.82,83 Grundläggande fibroblasttillväxtfaktor (eller FGF-2) är en medlem av fibroblasttillväxtfaktorfamiljen och förekommer högre i deprimerade än kontrollgrupper.84 Rapporterna är dock inte konsekventa; man fann att detta protein var lägre i MDD än friska kontroller, men minskade ytterligare tillsammans med antidepressiv behandling.85
Ytterligare tillväxtfaktorer som inte har undersökts tillräckligt vid depression inkluderar tyrosinkinas 2 och lösligt fms-liknande tyrosinkinas-1 (även kallat sVEGFR-1) som verkar i synergi med VEGF, och tyrosinkinasreceptorer (som binder BDNF) kan försvagas vid depression.86 Placental tillväxtfaktor är också en del av VEGF-familjen, men har såvitt vi vet inte studerats i systematiskt deprimerade prover.
Metaboliska biomarkörer vid depression
De viktigaste biomarkörerna förknippade med metabola sjukdomar inkluderar leptin, adiponektin, ghrelin, triglycerider, high-density lipoprotein (HDL), glukos, insulin och albumin.87 Sambanden mellan många av dessa och depression har granskats: leptin88 och ghrelin89 verkar lägre vid depression. än kontroller i periferin och kan öka tillsammans med antidepressiv behandling eller remission. Insulinresistens kan öka vid depression, om än i små mängder.90 Lipidprofiler, inklusive HDL-kolesterol, förefaller förändrade hos många patienter med depression, inklusive de utan samtidig fysisk sjukdom, även om detta förhållande är komplext och kräver ytterligare förklaring.91 Dessutom, hyperglykemi92 och hypoalbuminemi93 vid depression har rapporterats i recensioner.
Undersökningar av övergripande metabola tillstånd blir allt vanligare med hjälp av metabolomikpaneler av små molekyler i hopp om att hitta en robust biokemisk signatur för psykiatriska störningar. I en nyligen genomförd studie med modellering av artificiell intelligens var en uppsättning metaboliter som illustrerar ökad glukos-lipidsignalering mycket förutsägande för en MDD-diagnos,94 som stödde tidigare studier.95
Neurotransmittorfynd vid depression
Även om uppmärksamheten på monoaminer vid depression har gett relativt framgångsrika behandlingar, har inga robusta signalsubstansmarkörer identifierats för att optimera behandlingen baserat på selektiviteten hos monoaminmål för antidepressiva medel. Nyligen arbete pekar mot serotonin (5-hydroxitryptamin) 1A-receptorn som potentiellt viktig för både diagnos och prognos av depression, i väntan på nya genetiska och avbildningstekniker.96 Det finns nya potentiella behandlingar riktade mot 5-hydroxitryptamin; till exempel genom att använda en administrering med långsam frisättning av 5-hydroxitryptofan.97 Ökad överföring av dopamin interagerar med andra signalsubstanser för att förbättra kognitiva resultat såsom beslutsfattande och motivation.98 På samma sätt kan signalsubstanserna glutamat, noradrenalin, histamin och serotonin interagera och aktiveras som en del av en depressionsrelaterad stressreaktion; detta kan minska produktionen av 5-hydroxitryptamin genom �översvämning�. En nyligen genomförd granskning anger denna teori och föreslår att detta i TRD skulle kunna vändas (och 5-HT återställas) genom multimodal behandling riktad mot flera neurotransmittorer.99 Intressant nog sker ökningar av serotonin inte alltid tillsammans med terapeutiska antidepressiva fördelar.100 Trots detta , neurotransmittormetaboliter som 3-metoxi-4-hydroxifenylglykol, av noradrenalin eller homovanillinsyra av dopamin, har ofta visat sig öka tillsammans med minskning av depression med antidepressiv behandling101,102 eller att låga nivåer av dessa metaboliter förutsäger ett bättre svar på SSRI-behandling.102,103
Neuroendokrina fynd vid depression
Kortisol är den vanligaste biomarkören för HPA-axeln som har studerats vid depression. Många recensioner har fokuserat på de olika bedömningarna av HPA-aktivitet; totalt sett tyder dessa på att depression är associerad med hyperkortisolemi och att kortisoluppvaknandet ofta dämpas.104,105 Detta stöds av en nyligen genomförd granskning av kroniska kortisolnivåer uppmätt i hår, vilket stöder hypotesen om kortisolhyperaktivitet vid depression men hypoaktivitet vid andra sjukdomar, t.ex. som panikstörning.106 Dessutom kan särskilt förhöjda kortisolnivåer förutsäga ett sämre svar på psykologisk107 och antidepressiv108 behandling. Historiskt sett har den mest lovande neuroendokrina markören för prospektiv behandlingssvar varit dexametasonsuppressionstestet, där kortisol icke-suppression efter administrering av dexametason är associerad med en lägre sannolikhet för efterföljande remission. Detta fenomen har dock inte ansetts vara tillräckligt robust för klinisk tillämpning. Besläktade markörer kortikotrofinfrisättande hormon och adrenokortikotropinhormon samt vasopressin visar sig inkonsekvent överproduceras vid depression och dehydroepiandrosteron har visat sig vara försvagat; förhållandet mellan kortisol och dehydroepiandrosteron kan vara förhöjt som en relativt stabil markör i TRD, som kvarstår efter remission.109 Neuroendokrina hormondysfunktioner har länge varit förknippade med depression, och hypotyreos kan också spela en orsaksroll vid nedstämdhet.110 Vidare kan sköldkörtelreaktioner normalisera med framgångsrik behandling för depression.111
Inom ovanstående är det viktigt att även överväga signalvägar över system, såsom glykogensyntaskinas-3, mitogenaktiverat proteinkinas och cykliskt adenosin 3?,5?-monofosfat, involverat i synaptisk plasticitet112 och modifierat av antidepressiva medel.113 Ytterligare potentiella biomarkörkandidater som spänner över biologiska system i synnerhet mäts med hjälp av neuroimaging eller genetik. Som svar på bristen på robusta och meningsfulla genomiska skillnader mellan deprimerade och icke-deprimerade populationer, kan 114 nya genetiska tillvägagångssätt som polygena poäng115 eller telomerlängd116,117 visa sig vara mer användbara. Ytterligare biomarkörer som vinner popularitet är att undersöka cirkadiska cykler eller kronobiologiska biomarkörer som använder olika källor. Aktigrafi kan ge en objektiv bedömning av sömn- och vakenaktivitet och vila genom en accelerometer, och aktigrafiska enheter kan i allt högre grad mäta ytterligare faktorer som exponering för ljus. Detta kan vara mer användbart för upptäckt än vanliga subjektiva rapporter om patienter och skulle kunna ge nya prediktorer för behandlingssvar.118 Frågan om vilka biomarkörer som är mest lovande för translationell användning är en utmanande fråga, som utvidgas nedan.
Nuvarande utmaningar
För vart och ett av dessa fem neurobiologiska system som granskas följer bevisen en liknande berättelse: det finns många biomarkörer som finns som i vissa avseenden är förknippade med depression. Dessa markörer är ofta relaterade till varandra på ett komplext sätt som är svårt att modellera. Bevisen är inkonsekvent, och det är troligt att vissa är epifenomen av andra faktorer och vissa är viktiga för endast en delmängd av patienter. Biomarkörer kommer sannolikt att vara användbara genom en mängd olika vägar (t.ex. de som förutsäger efterföljande svar på behandlingen, de som indikerar att specifika behandlingar är mer sannolikt att vara effektiva eller de som förändras med interventioner oavsett kliniska förbättringar). Nya metoder krävs för att maximera konsekvens och klinisk användbarhet av biologiska bedömningar i psykiatriska populationer.
Biomarkörvariabilitet
Variation av biomarkörer över tid och över situationer hänför sig mer till vissa typer (t.ex. proteomik) än andra (genomik). Standardiserade normer för många finns inte eller har inte blivit allmänt accepterade. Faktum är att miljöfaktorers inverkan på markörer ofta beror på genetisk sammansättning och andra fysiologiska skillnader mellan människor som inte alla kan förklaras. Detta gör bedömningen av biomarköraktivitet och identifiering av biologiska avvikelser svårtolkad. På grund av antalet potentiella biomarkörer har många inte mätts brett eller i en komplett panel tillsammans med andra relevanta markörer.
Många faktorer har rapporterats förändra proteinnivåerna över biologiska system hos patienter med affektiva störningar. Tillsammans med forskningsrelaterade faktorer såsom varaktighet och förhållanden för lagring (som kan orsaka nedbrytning av vissa föreningar), inkluderar dessa tid på dygnet uppmätt, etnicitet, träning,119 diet (t.ex. mikrobiomaktivitet, särskilt förutsatt att de flesta blodbiomarkörstudier gör det kräver inte ett fasteprov),120 rökning och droganvändning,121 samt hälsofaktorer (såsom komorbida inflammatoriska, kardiovaskulära eller andra fysiska sjukdomar). Till exempel, även om förhöjd inflammation observeras hos deprimerade men i övrigt friska individer jämfört med icke-deprimerade grupper, har deprimerade individer som också har ett komorbidt immunrelaterat tillstånd ofta ännu högre nivåer av cytokiner än de utan depression eller sjukdom.122 Vissa framträdande faktorer med trolig inblandning i sambandet mellan biomarkörer, depression och behandlingssvar beskrivs nedan.
Stress. Både endokrina och immunsvar har välkända roller för att svara på stress (fysiologisk eller psykologisk), och övergående stress vid tidpunkten för biologisk provtagning mäts sällan i forskningsstudier trots variationen av denna faktor mellan individer som kan accentueras av nuvarande depressiva symtom. Både akuta och kroniska psykologiska stressorer fungerar som en immunutmaning, och accentuerar inflammatoriska svar på kort och längre sikt.123,124 Detta fynd sträcker sig till upplevelsen av stress i tidigt liv, som har associerats med vuxna inflammatoriska förhöjningar som är oberoende av stress som upplevs som en vuxen.125,126 127 Under barndomens traumatiska erfarenheter har förhöjd inflammation också rapporterats endast hos de barn som för närvarande var deprimerade.128 Omvänt kan personer med depression och en historia av barndomstrauma ha avtrubbade kortisolsvar på stress, jämfört med de med depression och inget trauma i tidigt liv.129 Stressinducerade förändringar i HPA-axeln verkar vara relaterade till kognitiv funktion,130 såväl som depressionssubtyp eller variation i HPA-relaterade gener.131 Stress har också kort- och långtidsförsämrande effekter på neurogenes132 och andra neurala nerver. mekanismer.XNUMX Det är oklart exakt hur barndomstrauma påverkar biologiska markörer hos deprimerade vuxna s, men det är möjligt att stress tidigt i livet predisponerar vissa individer för att uthärda stressreaktioner i vuxen ålder som förstärks psykologiskt och/eller biologiskt.
Kognitiv funktion. Neurokognitiva dysfunktioner förekommer ofta hos personer med affektiva störningar, även vid omedicinerad MDD.133 Kognitiva brister verkar kumulativa tillsammans med behandlingsresistens.134 Neurobiologiskt kommer HPA-axeln129 och neurotrofiska system135 sannolikt att spela en nyckelroll i detta förhållande. Neurotransmittorer noradrenalin och dopamin är sannolikt viktiga för kognitiva processer som inlärning och minne.136 Förhöjda inflammatoriska svar har kopplats till kognitiv försämring och påverkar sannolikt kognitiv funktion i depressiva episoder137 och vid remission, genom en mängd olika mekanismer.138 Ja, Krogh et al139 föreslog att CRP är närmare relaterat till kognitiv prestation än till kärnsymtomen på depression.
Ålder, kön och BMI. Frånvaron eller närvaron och riktningen av biologiska skillnader mellan män och kvinnor har varit särskilt varierande i bevisen hittills. Neuroendokrina hormonvariationer mellan män och kvinnor interagerar med depressionskänslighet.140 En genomgång av inflammationsstudier rapporterade att kontroll för ålder och kön inte påverkade patientkontrollskillnader i inflammatoriska cytokiner (även om sambandet mellan IL-6 och depression minskade när åldern ökade, vilket stämmer överens med teorier om att inflammation generellt ökar med åldern).41,141 77 VEGF-skillnader mellan patienter och kontroller är större i studier som bedömer yngre prov, medan kön, BMI och kliniska faktorer inte påverkade dessa jämförelser på en metaanalytisk nivå.41 bristen på justering för BMI i tidigare undersökningar av inflammation och depression tycks förvirra mycket signifikanta skillnader som rapporterats mellan dessa grupper.142 Förstorad fettvävnad har definitivt visat sig stimulera cytokinproduktionen samt vara nära kopplad till metabola markörer.XNUMX Eftersom psykotropa läkemedel kan förknippas med wei ght ökning och ett högre BMI, och dessa har associerats med behandlingsresistens vid depression, är detta ett viktigt område att undersöka.
Medicinering. Många biomarkörstudier vid depression (både tvärsnitts- och longitudinella) har samlat in baslinjeprover hos icke-medicinerade deltagare för att minska heterogeniteten. Men många av dessa bedömningar görs efter en uttvättningsperiod från medicinering, vilket lämnar den potentiellt betydande förvirrande faktorn av kvarvarande förändringar i fysiologi, förvärrad av det omfattande utbud av behandlingar som finns tillgängliga som kan ha haft olika effekter på inflammation. Vissa studier har uteslutit psykotropa, men inte andra läkemedelsanvändningar: i synnerhet är p-piller ofta tillåtet hos forskningsdeltagare och kontrolleras inte för i analyser, vilket nyligen har indikerats öka hormon- och cytokinnivåerna.143,144 Flera studier indikerar att antidepressiva medel mediciner har effekter på det inflammatoriska svaret,34,43,49,145�147 HPA-axeln,108 neurotransmittor,148 och neurotrofisk149 aktivitet. De många potentiella behandlingarna för depression har dock distinkta och komplexa farmakologiska egenskaper, vilket tyder på att det kan finnas diskreta biologiska effekter av olika behandlingsalternativ, vilket stöds av aktuella data. Det har teoretiserats att förutom monoamineffekter kommer specifika serotonininriktade läkemedel (dvs. SSRI) sannolikt att inrikta sig på Th2-förskjutningar i inflammation, och noradrenerga antidepressiva medel (t.ex. SNRI) påverkar ett Th1-skifte.150 Det är ännu inte möjligt att bestämma effekterna av individuella eller kombinationsläkemedel på biomarkörer. Dessa förmedlas sannolikt av andra faktorer, inklusive behandlingens längd (få prövningar bedömer långvarig läkemedelsanvändning), provets heterogenitet och att deltagarna inte stratifieras efter behandlingssvar.
Heterogenitet
Metodologiska. Som antytts ovan är skillnader (mellan och inom studier) när det gäller vilka behandlingar (och kombinationer) deltagarna tar och har tagit tidigare skyldiga att introducera heterogenitet i forskningsresultat, särskilt inom biomarkörforskning. Utöver detta varierar många andra design- och provegenskaper mellan studierna, vilket ökar svårigheten med att tolka och tillskriva fynd. Dessa inkluderar mätparametrar för biomarkörer (t.ex. analyskit) och metoder för att samla in, lagra, bearbeta och analysera markörer vid depression. Hiles et al141 undersökte några källor till inkonsekvens i litteraturen om inflammation och fann att noggrannheten av depressionsdiagnos, BMI och komorbida sjukdomar var viktigast att ta hänsyn till vid bedömning av perifer inflammation mellan deprimerade och icke-deprimerade grupper.
Klinisk. Den omfattande heterogeniteten hos deprimerade populationer är väl dokumenterad151 och är en avgörande bidragande orsak till kontrasterande fynd inom forskningslitteraturen. Det är troligt att även inom diagnoser är onormala biologiska profiler begränsade till undergrupper av individer som kanske inte är stabila över tid. Sammanhållna undergrupper av personer som lider av depression kan identifieras genom en kombination av psykologiska och biologiska faktorer. Nedan skisserar vi potentialen för att utforska undergrupper för att möta de utmaningar som biomarkörernas variation och heterogenitet utgör.
Subtyper inom depression
Hittills har inga homogena undergrupper inom depressionsepisoder eller störningar på ett tillförlitligt sätt kunnat skilja mellan patienter baserat på symtompresentationer eller behandlingslyhördhet.152 Förekomsten av en undergrupp där biologiska aberrationer är mer uttalade skulle bidra till att förklara heterogeniteten mellan tidigare studier och kan katalysera vägen mot stratifierad behandling. Kunugi et al153 har föreslagit en uppsättning av fyra potentiella subtyper baserade på rollen av olika neurobiologiska system som uppvisar kliniskt relevanta subtyper i depression: de med hyperkortisolism som presenterar melankolisk depression, eller hypokortisolism som speglar en atypisk subtyp, en dopaminrelaterad undergrupp av patienter som kan förekommer framträdande med anhedoni (och skulle kunna svara bra på t.ex. aripiprazol) och en inflammatorisk subtyp som kännetecknas av förhöjd inflammation. Många artiklar som fokuserar på inflammation har specificerat fallet för förekomsten av en "inflammatorisk subtyp" inom depression.55,56,154,155 Kliniska korrelat av förhöjd inflammation är ännu obestämda och få direkta försök har gjorts för att upptäcka vilka deltagare som kan utgöra denna kohort. Det har föreslagits att personer med atypisk depression kan ha högre nivåer av inflammation än den melankoliska subtypen,156 vilket kanske inte stämmer överens med fynden angående HPA-axeln vid melankoliska och atypiska subtyper av depression. TRD37 eller depression med framträdande somatiska symtom157 har också ansetts vara en potentiell inflammatorisk subtyp, men neurovegetativa (sömn, aptit, förlust av libido), humör (inklusive nedstämdhet, suicidalitet och irritabilitet) och kognitiva symtom (inklusive affektiv fördom och skuld)158 alla verkar relaterade till biologiska profiler. Ytterligare potentiella kandidater för en inflammatorisk subtyp involverar upplevelsen av sjukdomsbeteendeliknande symtom159,160 eller ett metabolt syndrom.158
Benägenheten till (hypo)mani kan skilja biologiskt mellan patienter som lider av depression. Bevis tyder nu på att bipolära sjukdomar är en mångfacetterad grupp av humörstörningar, där bipolär subsyndromal störning förekommer mer utbrett än vad som tidigare erkänts.161 Felaktig och/eller fördröjd upptäckt av bipolär sjukdom har nyligen framhållits som ett stort problem inom klinisk psykiatri, med Den genomsnittliga tiden för att korrigera diagnosen överskrider ofta ett decennium162 och denna fördröjning orsakar större svårighetsgrad och kostnaden för den totala sjukdomen.163 Med majoriteten av patienter med bipolär sjukdom som initialt uppvisar en eller flera depressiva episoder och unipolär depression är den vanligaste feldiagnostiken, identifiering av faktorer som kan skilja mellan unipolär och bipolär depression har betydande implikationer.164 Bipolära spektrumstörningar har sannolikt inte upptäckts i några tidigare MDD-biomarkörundersökningar, och bristfälliga bevis har indikerat differentiering av HPA-axelaktivitet109 eller inflammation165,166 mellan bipolär och unipolär depr session. Dessa jämförelser är dock få, har små urvalsstorlekar, identifierade icke-signifikanta trendeffekter eller rekryterade populationer som inte var väl karakteriserade av diagnos. Dessa undersökningar undersöker inte heller vilken roll behandlingslyhördhet spelar i dessa relationer.
Både bipolära sjukdomar167 och behandlingsresistens168 är inte dikotoma konstruktioner och ligger på kontinuitet, vilket ökar utmaningen med subtypidentifiering. Förutom subtypning är det värt att notera att många biologiska avvikelser som observerats vid depression på liknande sätt återfinns hos patienter med andra diagnoser. Därmed är även transdiagnostiska undersökningar potentiellt viktiga.
Utmaningar för mätning av biomarkörer
Val av biomarkörer. Det stora antalet potentiellt användbara biomarkörer utgör en utmaning för psykobiologin när det gäller att avgöra vilka markörer som är inblandade på vilket sätt och för vem. För att öka utmaningen har relativt få av dessa biomarkörer varit föremål för tillräcklig undersökning vid depression, och för de flesta är deras exakta roller i friska och kliniska populationer inte väl förstått. Trots detta har ett antal försök gjorts att föreslå lovande biomarkörpaneler. Förutom Brand et al:s 16 uppsättningar av markörer med stark potential27, beskriver Lopresti et al ytterligare en omfattande uppsättning markörer för oxidativ stress med potential för att förbättra behandlingssvaret.28 Papakostas et al definierade a priori en uppsättning av nio serummarkörer som spänner över biologiska system (BDNF, kortisol, löslig TNFa-receptor typ II, alfal-antitrypsin, apolipoprotein CIII, epidermal tillväxtfaktor, myeloperoxidas, prolaktin och resistin) i validerings- och replikationsprover med MDD. När de väl kombinerats kunde ett sammansatt mått på dessa nivåer skilja mellan MDD och kontrollgrupper med 1 %–80 % noggrannhet.90 Vi föreslår att inte ens dessa täcker alla potentiella kandidater inom detta område; se tabell 169 för en icke uttömmande beskrivning av biomarkörer med potential för depression, innehållande både de med en evidensbas och lovande nya markörer.
Teknologi. På grund av tekniska framsteg är det nu möjligt (verkligen bekvämt) att mäta ett stort utbud av biomarkörer samtidigt till en lägre kostnad och med högre känslighet än vad som varit fallet tidigare. För närvarande är denna förmåga att mäta många föreningar före vår förmåga att effektivt analysera och tolka data,170 något som kommer att fortsätta med ökningen av biomarkörer och nya markörer som med metabolomik. Detta beror till stor del på en bristande förståelse för de exakta rollerna för och de inbördes relationerna mellan markörer, och ett otillräckligt grepp om hur relaterade markörer associerar över olika biologiska nivåer (t.ex. genetisk, transkription, protein) inom och mellan individer. Big data som använder nya analytiska tillvägagångssätt och standarder kommer att hjälpa till att hantera detta, och nya metoder föreslås; ett exempel är utvecklingen av ett statistiskt tillvägagångssätt grundat på flödesbaserad analys för att upptäcka nya potentiella metaboliska markörer baserade på deras reaktioner mellan nätverk och integrera genuttryck med metabolitdata.171 Maskininlärningstekniker tillämpas redan och kommer att hjälpa till med modeller som använder biomarkörer data för att förutsäga behandlingsresultat i studier med stora data.172
Aggregerande biomarkörer. Att undersöka en rad biomarkörer samtidigt är ett alternativ till att inspektera isolerade markörer som skulle kunna ge en mer exakt syn på den komplexa väven av biologiska system eller nätverk.26 Dessutom för att hjälpa till med att reda ut kontrasterande bevis i denna litteratur hittills (särskilt där biomarkörnätverk och interaktioner är väl förstådda), kan biomarkördata sedan aggregeras eller indexeras. En utmaning är att identifiera den optimala metoden för att genomföra detta, och det kan kräva förbättringar av teknik och/eller nya analytiska tekniker (se avsnittet "Big data"). Historiskt sett har förhållanden mellan två distinkta biomarkörer gett intressanta fynd.109,173 174 Få försök har gjorts att aggregera biomarkördata i större skala, som de som använder principal komponentanalys av proinflammatoriska cytokinnätverk.43 I en metaanalys har proinflammatoriska cytokiner varit omvandlas till en enstaka effektstorlekspoäng för varje studie, och visade totalt sett signifikant högre inflammation före antidepressiv behandling, vilket förutsäger efterföljande bortfall i öppenvårdsstudier. Sammansatta biomarkörpaneler är både en utmaning och möjlighet för framtida forskning för att identifiera meningsfulla och tillförlitliga resultat som kan användas för att förbättra behandlingsresultat.80 En studie av Papakostas et al tog ett alternativt tillvägagångssätt och valde en panel av heterogena serumbiomarkörer (av inflammatoriska, HPA-axel och metabola system) som hade indikerats skilja sig mellan deprimerade och kontrollpersoner i en tidigare studie och sammansatte dessa till ett riskpoäng som skilde sig i två oberoende prover och en kontrollgrupp med >169 % sensitivitet och specificitet.XNUMX
Stora data. Användningen av big data är förmodligen nödvändig för att ta itu med de nuvarande utmaningarna som beskrivs kring heterogenitet, biomarkörvariabilitet, identifiera de optimala markörerna och föra fältet mot translationell, tillämpad forskning inom depression. Men, som beskrivits ovan, medför detta tekniska och vetenskapliga utmaningar.175 Hälsovetenskapen har först nyligen börjat använda big data-analys, något decennium senare än i näringslivet. Studier som iSPOT-D152 och konsortier som Psychiatric Genetics Consortium176 går dock framåt med vår förståelse av biologiska mekanismer inom psykiatrin. Maskininlärningsalgoritmer har i väldigt få studier börjat tillämpas på biomarkörer för depression: en nyligen genomförd undersökning samlade data från >5,000 250 deltagare av 21 biomarkörer; efter multipel imputering av data genomfördes en maskininlärningsförstärkt regression, vilket indikerar 177 potentiella biomarkörer. Efter ytterligare regressionsanalyser valdes tre biomarkörer ut som starkast associerade med depressiva symtom (mycket varierande röda blodkroppar, serumglukos och bilirubinnivåer). Författarna drar slutsatsen att big data kan användas effektivt för att generera hypoteser.XNUMX Större biomarkörfenotypningsprojekt pågår nu och kommer att bidra till att främja vår resa in i framtiden för depressionens neurobiologi.
Framtida prospekt
Identifiering av biomarkörpanel
Fynden i litteraturen hittills kräver replikering i storskaliga studier. Detta gäller särskilt för nya biomarkörer, såsom kemokin tymus och aktiveringsreglerad kemokin och tillväxtfaktorn tyrosinkinas 2 som, såvitt vi vet, inte har undersökts i kliniskt deprimerade och friska kontrollprover. Big data-studier måste analysera omfattande biomarkörpaneler och använda sofistikerade analystekniker för att fullt ut fastställa sambanden mellan markörer och de faktorer som modifierar dem i kliniska och icke-kliniska populationer. Dessutom kan storskaliga replikeringar av huvudkomponentanalys etablera starkt korrelerade grupper av biomarkörer och kan också informera om användningen av "kompositer" i biologisk psykiatri, vilket kan förbättra homogeniteten i framtida fynd.
Upptäckt av homogena subtyper
När det gäller urval av biomarkörer kan flera paneler krävas för olika potentiella vägar som forskning kan implicera. Sammantaget indikerar de aktuella bevisen att biomarkörprofiler säkert, men kraftigt förändras i en subpopulation av individer som för närvarande lider av depression. Detta kan fastställas inom eller över diagnostiska kategorier, vilket skulle förklara en viss inkonsekvens av fynd som kan observeras i denna litteratur. Att kvantifiera en biologisk undergrupp (eller undergrupper) kan mest effektivt underlättas av en stor klusteranalys av biomarkörnätverkspaneler i depression. Detta skulle illustrera variation inom populationen; latenta klassanalyser skulle kunna uppvisa distinkta kliniska egenskaper baserade på till exempel inflammation.
Specifika behandlingseffekter på inflammation och respons
Alla vanligt föreskrivna behandlingar för depression bör utvärderas heltäckande för deras specifika biologiska effekter, även ta hänsyn till effektiviteten av behandlingsförsök. Detta kan göra det möjligt för konstruktioner relaterade till biomarkörer och symtompresentationer att förutsäga resultat av en mängd olika antidepressiva behandlingar på ett mer personligt sätt, och kan vara möjligt i samband med både unipolär och bipolär depression. Detta kommer sannolikt att vara användbart för nya potentiella behandlingar såväl som för närvarande indikerade behandlingar.
Prospektiv bestämning av behandlingssvar
Användning av ovanstående tekniker kommer sannolikt att resultera i en förbättrad förmåga att prospektivt förutse behandlingsresistens. Mer autentiska och ihållande (t.ex. långsiktiga) mått på behandlingssvar kan bidra till detta. Bedömning av andra giltiga mått på patientens välbefinnande (såsom livskvalitet och vardagsfunktion) skulle kunna ge en mer holistisk bedömning av behandlingsresultat som kan associeras närmare med biomarkörer. Även om biologisk aktivitet ensam kanske inte kan skilja behandlingssvarare från icke-svarare, kan samtidig mätning av biomarkörer med psykosociala eller demografiska variabler integreras med biomarkörinformation för att utveckla en prediktiv modell för otillräckligt behandlingssvar. Om en tillförlitlig modell utvecklas för att förutsäga respons (antingen för den deprimerade befolkningen eller en subpopulation) och valideras retrospektivt, kan en translationell design fastställa dess tillämpbarhet i en stor kontrollerad studie.
Mot stratifierade behandlingar
För närvarande är patienter med depression inte systematiskt inriktade på att få ett optimerat interventionsprogram. Om den är validerad kan en stratifierad studiedesign användas för att testa en modell för att förutsäga bortfall och/eller för att bestämma var en patient behöver triageras i en stegvis vårdmodell. Detta kan vara användbart i både standardiserade och naturalistiska behandlingsmiljöer, över olika typer av interventioner. I slutändan kan en kliniskt genomförbar modell utvecklas för att ge individer den mest lämpliga behandlingen, för att känna igen dem som sannolikt kommer att utveckla refraktär depression och ge förbättrad vård och övervakning till dessa patienter. Patienter som identifieras som risk för behandlingsresistens kan ordineras en samtidig psykologisk och farmakologisk behandling eller kombinationsfarmakoterapi. Som ett spekulativt exempel kan deltagare utan proinflammatoriska cytokinförhöjningar indikeras att få psykologisk snarare än farmakologisk terapi, medan en undergrupp av patienter med särskilt hög inflammation kan få ett antiinflammatoriskt medel som en förstärkning till standardbehandling. I likhet med stratifiering kan personliga behandlingsvalsstrategier vara möjliga i framtiden. Till exempel kan en viss deprimerad individ ha markant hög TNF? nivåer, men inga andra biologiska avvikelser, och skulle kunna dra nytta av korttidsbehandling med en TNF? antagonist.54 Personlig behandling kan också innebära övervakning av uttryck av biomarkörer under behandling för att informera om möjliga interventionsförändringar, längden på fortsatt behandling som krävs eller för att upptäcka tidiga markörer för återfall.
Nya behandlingsmål
Det finns ett stort antal potentiella behandlingar som kan vara effektiva för depression, som inte har undersökts tillräckligt, inklusive nya eller återanvända interventioner från andra medicinska discipliner. Några av de mest populära målen har varit i antiinflammatoriska läkemedel som celecoxib (och andra cyklooxygenas-2-hämmare), TNF? antagonister etanercept och infliximab, minocyklin eller aspirin. Dessa verkar lovande.178 Antiglukokortikoidföreningar, inklusive ketokonazol179 och metyrapon,180 har undersökts för depression, men båda har nackdelar med sin biverkningsprofil och den kliniska potentialen för metyrapon är osäker. Mifepriston181 och kortikosteroiderna fludrokortison och spironolakton,182 och dexametason och hydrokortison183 kan också vara effektiva vid behandling av depression på kort sikt. Inriktning mot glutamat-N-metyl-d-aspartatreceptorantagonister, inklusive ketamin, kan representera effektiva behandlingar vid depression.184 Omega-3 fleromättade fettsyror påverkar inflammatorisk och metabolisk aktivitet och verkar visa viss effektivitet för depression.185 Det är möjligt att statiner kan har antidepressiva effekter186 genom relevanta neurobiologiska vägar.187
På detta sätt har de biokemiska effekterna av antidepressiva medel (se avsnittet �Medicinering�) utnyttjats för kliniska fördelar inom andra discipliner: särskilt gastroenterologiska, neurologiska och ospecifika symtomsjukdomar.188 Antiinflammatoriska effekter av antidepressiva läkemedel kan utgöra en del av mekanismen för dessa förmåner. Litium har också föreslagits minska inflammation, kritiskt genom glykogensyntaskinas-3-vägar.189 En fokusering på dessa effekter kan visa sig vara informativ för en depressionsbiomarkörsignatur och i sin tur kan biomarkörer representera surrogatmarkörer för utveckling av nya läkemedel.
Dr Alex Jimenez insikt
Depression är en psykisk störning som kännetecknas av allvarliga symtom som påverkar humöret, inklusive förlust av intresse för aktiviteter. Nya forskningsstudier har dock funnit att det kan vara möjligt att diagnostisera depression med mer än bara patientens beteendesymtom. Enligt forskarna är identifiering av lättillgängliga biomarkörer som mer exakt kan diagnostisera depression grundläggande för att förbättra en patients allmänna hälsa och välbefinnande. Till exempel tyder kliniska fynd på att individer med egentlig depression, eller MDD, har lägre nivåer av molekylen acetyl-L-karnitin, eller LAC, i blodet än friska kontroller. I slutändan kan fastställande av biomarkörer för depression potentiellt hjälpa till att bättre avgöra vem som löper risk att utveckla sjukdomen samt hjälpa vårdpersonal att avgöra det bästa behandlingsalternativet för en patient med depression.
Slutsats
Litteraturen indikerar att cirka två tredjedelar av patienter med depression inte uppnår remission till en initial behandling och att sannolikheten för bortfall ökar med antalet prövade behandlingar. Att tillhandahålla ineffektiva terapier har betydande konsekvenser för individuella och samhälleliga kostnader, inklusive ihållande nöd och dåligt välbefinnande, risk för självmord, förlust av produktivitet och slöseri med sjukvårdsresurser. Den omfattande litteraturen om depression indikerar ett stort antal biomarkörer med potential att förbättra behandlingen för personer med depression. Förutom signalsubstanser och neuroendokrina markörer som har varit föremål för omfattande studier under många decennier, framhäver nya insikter det inflammatoriska svaret (och immunsystemet mer generellt), metabola faktorer och tillväxtfaktorer som är viktiga för depression. Men överdrivet kontrasterande bevis illustrerar att det finns ett antal utmaningar som måste hanteras innan biomarkörforskning kan tillämpas för att förbättra hanteringen och vården av personer med depression. På grund av den rena komplexiteten hos biologiska system är samtidiga undersökningar av ett omfattande utbud av markörer i stora prover avsevärda fördelar för att upptäcka interaktioner mellan biologiska och psykologiska tillstånd mellan individer. Att optimera mätningen av både neurobiologiska parametrar och kliniska mått på depression kommer sannolikt att underlätta större förståelse. Denna översyn belyser också vikten av att undersöka potentiellt modifierande faktorer (såsom sjukdom, ålder, kognition och medicinering) för att få en sammanhängande förståelse av depressionens biologi och mekanismer för behandlingsresistens. Det är troligt att vissa markörer kommer att visa mest lovande för att förutsäga behandlingssvar eller motstånd mot specifika behandlingar i en undergrupp av patienter, och samtidig mätning av biologiska och psykologiska data kan förbättra förmågan att prospektivt identifiera de som löper risk för dåliga behandlingsresultat. Att inrätta en biomarkörpanel har konsekvenser för att öka diagnostisk noggrannhet och prognos, såväl som för att individualisera behandlingar vid det tidigaste möjliga skedet av depressiv sjukdom och utveckla effektiva nya behandlingsmål. Dessa implikationer kan vara begränsade till undergrupper av deprimerade patienter. Vägarna mot dessa möjligheter kompletterar nyare forskningsstrategier för att koppla kliniska syndrom närmare till underliggande neurobiologiska substrat.6 Förutom att minska heterogeniteten kan detta underlätta en förskjutning mot paritet i aktning mellan fysisk och mental hälsa. Det är tydligt att även om det krävs mycket arbete, har upprättandet av sambandet mellan relevanta biomarkörer och depressiva störningar betydande konsekvenser för att minska bördan av depression på individuell och samhällelig nivå.
Erkännanden
Den här rapporten representerar oberoende forskning finansierad av National Institute for Health Research (NIHR) Biomedical Research Centre vid South London och Maudsley NHS Foundation Trust och King's College London. Åsikterna som uttrycks är författarnas och inte nödvändigtvis de från NHS, NIHR eller Department of Health.
fotnoter
Avslöjande. AHY har under de senaste 3 åren fått hedersbetygelser för att tala av Astra Zeneca (AZ), Lundbeck, Eli Lilly, Sunovion; hedersbetygelse för konsulttjänster från Allergan, Livanova och Lundbeck, Sunovion, Janssen; och forskningsbidrag från Janssen och brittiska finansieringsorgan (NIHR, MRC, Wellcome Trust). AJC har under de senaste 3 åren fått hedersbetygelser för att tala från Astra Zeneca (AZ), hedersbetygelser för konsultation från Allergan, Livanova och Lundbeck, och forskningsanslagsstöd från Lundbecks och brittiska finansieringsorgan (NIHR, MRC, Wellcome Trust).
Författarna rapporterar inga andra intressekonflikter i detta arbete.
SammanfattningsvisÄven om många forskningsstudier har hittat hundratals biomarkörer för depression, har inte många fastställt sin roll i depressiv sjukdom eller hur exakt biologisk information kan användas för att förbättra diagnos, behandling och prognos. Men artikeln ovan granskar den tillgängliga litteraturen om biomarkörerna som är involverade under andra processer och jämför de kliniska fynden med depression. Dessutom kan nya rön om biomarkörer för depression hjälpa till att bättre diagnostisera depression för att följa upp med bättre behandling. Information som refereras från National Center for Biotechnology Information (NCBI).�Omfattningen av vår information är begränsad till kiropraktik såväl som till ryggradsskador och tillstånd. För att diskutera ämnet är du välkommen att fråga Dr Jimenez eller kontakta oss på�915-850-0900 .
Curated av Dr. Alex Jimenez
Ytterligare ämnen: Ryggsmärta
Ryggont är en av de vanligaste orsakerna till funktionshinder och missade dagar på jobbet över hela världen. Faktum är att ryggsmärta har tillskrivits som den näst vanligaste anledningen till doktorsbesök, som endast överträffas av övre luftvägsinfektioner. Cirka 80 procent av befolkningen kommer att uppleva någon typ av ryggsmärta minst en gång i livet. Ryggraden är en komplex struktur bestående av ben, leder, ligament och muskler, bland annat mjuka vävnader. På grund av detta, skador och / eller förvärrade förhållanden, till exempel hernierade skivor, kan så småningom leda till symtom på ryggont. Sportskador eller skador på bilolyckor är ofta den vanligaste orsaken till ryggsmärta, men ibland kan de enklaste rörelserna få ont. Lyckligtvis kan alternativa behandlingsalternativ, som kiropraktikvård, hjälpa till att lindra ryggont genom användningen av ryggmärgsjusteringar och manuell manipulation, vilket i slutändan förbättrar smärtlindringen.
1.�Prince M, Patel V, Saxena S, et al. Ingen hälsa utan mental hälsa.�Lancet.�2007;370(9590):859�877.[PubMed]
2.�Kingdon D, Wykes T. Ökad finansiering behövs för forskning om mental hälsa.�BMJ.�2013;346:f402.[PubMed]
3.�Vivekanantham S, Strawbridge R, Rampuri R, Ragunathan T, Young AH. Publikationsparitet för psykiatri.�Br J Psykiatri.�2016;209(3):257�261.�[PubMed]
4.�Fava M. Diagnos och definition av behandlingsresistent depression.�Biol Psykiatri.�2003;53(8):649�659.�[PubMed]
5.�Insel T, Cuthbert B, Garvey M, et al. Forskningsdomänkriterier (RDoC): mot ett nytt klassificeringsramverk för forskning om psykiska störningar.�Am J Psychiatry.�2010;167(7):748�751.�[PubMed]
6.�Kapur S, Phillips AG, Insel TR. Varför har det tagit så lång tid för biologisk psykiatri att utveckla kliniska tester och vad man ska göra åt det.�Mol Psykiatri.�2012;17(12):1174�1179.�[PubMed]
7.�Gaynes BN, Warden D, Trivedi MH, Wisniewski SR, Fava M, Rush JA. Vad lärde STAR*D oss? Resultat från en storskalig, praktisk, klinisk prövning för patienter med depression.�Psychiatr Serv.�2009;60(11):1439�1445.�[PubMed]
8.�Fekadu A, Rane LJ, Wooderson SC, Markopoulou K, Poon L, Cleare AJ. Förutsägelse av långsiktiga resultat av behandlingsresistent depression inom tertiärvård.�Br J Psykiatri.�2012;201(5):369�375.[PubMed]
9.�Fekadu A, Wooderson SC, Markopoulo K, Donaldson C, Papadopoulos A, Cleare AJ. Vad händer med patienter med behandlingsresistent depression? En systematisk översikt av medellång till lång sikt resultatstudier.�J Affect Disord.�2009;116(1�2):4�11.�[PubMed]
10.�Trivedi M. Behandlingsstrategier för att förbättra och upprätthålla remission vid egentlig depression.�Dialogues Clin Neurosci.�2008;10(4): 377.�[PMC gratis artikel]�[PubMed]
11.�Fekadu A, Wooderson SC, Markopoulou K, Cleare AJ. Maudsley Staging-metoden för behandlingsresistent depression: förutsägelse av långsiktiga resultat och kvarstående symtom.�J Clin Psykiatri.�2009;70(7):952�957.�[PubMed]
12.�Bennabi D, Aouizerate B, El-Hage W, et al. Riskfaktorer för behandlingsresistens vid unipolär depression: en systematisk översikt.�J Affect Disord.�2015;171:137�141.�[PubMed]
13.�Serretti A, Olgiati P, Liebman MN, et al. Klinisk förutsägelse av antidepressivt svar vid humörstörningar: linjära multivariata kontra neurala nätverksmodeller.�Psychiatry Res.�2007;152(2�3):223�231.[PubMed]
14.�Driessen E, Hollon SD. Kognitiv beteendeterapi för humörstörningar: effekt, moderatorer och mediatorer.�Psykiatriker Clin North Am.�2010;33(3):537�555.�[PMC gratis artikel]�[PubMed]
15.�Cleare A, Pariante C, Young A, et al. Medlemmar av Consensus Meeting Evidensbaserade riktlinjer för behandling av depressiva sjukdomar med antidepressiva medel: en revidering av 2008 års brittiska föreningen för psykofarmakologi.�J Psychopharmacol.�2015;29(5):459�525.�[PubMed]
16.�Tunnard C, Rane LJ, Wooderson SC, et al. Effekten av motgångar i barndomen på suicidalitet och kliniskt förlopp vid behandlingsresistent depression.�J Affect Disord.�2014;152 154:122�130.�[PubMed]
17.�Nemeroff CB, Heim CM, Thase ME, et al. Differentiella svar på psykoterapi jämfört med farmakoterapi hos patienter med kroniska former av egentlig depression och barndomstrauma.�Proc Natl Acad Sci US A.�2003;100(24):14293�14296.�[PMC gratis artikel]�[PubMed]
18.�Nierenberg AA. Prediktorer för svar på antidepressiva allmänna principer och kliniska implikationer.�Psykiatriker Clin North Am.�2003;26(2):345�352.�[PubMed]
19.�Det är JAG. Använda biomarkörer för att förutsäga behandlingssvar vid allvarlig depressiv sjukdom: bevis från tidigare och nuvarande studier.�Dialogues Clin Neurosci.�2014;16(4):539�544.�[PMC gratis artikel]�[PubMed]
20.�Jani BD, McLean G, Nicholl BI, et al. Riskbedömning och förutsägelse av resultat hos patienter med depressiva symtom: en genomgång av den potentiella rollen av perifera blodbaserade biomarkörer.�Front Hum Neurosci.�2015;9:18.�[PMC gratis artikel]�[PubMed]
21.�Suravajhala P, Kogelman LJ, Kadarmideen HN. Multiomisk dataintegration och analys med hjälp av systemgenomik: metoder och tillämpningar inom djurproduktion, hälsa och välfärd.�Genet Sel Evol.�2016;48(1): 1.�[PMC gratis artikel]�[PubMed]
22.�Menke A. Genuttryck: Biomarkör för antidepressiv terapi?�Int Rev Psychiatry.�2013;25(5):579�591.�[PubMed]
23.�Peng B, Li H, Peng XX. Funktionell metabolomik: från upptäckt av biomarkörer till omprogrammering av metabolom.�Proteincell.�2015;6(9):628�637.�[PMC gratis artikel]�[PubMed]
24.�Aagaard K, Petrosino J, Keitel W, et al. Human Microbiome Project-strategin för omfattande provtagning av den mänskliga mikrobiomen och varför det är viktigt.�FASEB J.�2013;27(3):1012�1022.[PMC gratis artikel]�[PubMed]
25.�Sonner Z, Wilder E, Heikenfeld J, et al. Mikrofluidikerna i den ekrina svettkörteln, inklusive biomarköruppdelning, transport och biosensing implikationer.�Biomikrofluidik.�2015;9(3): 031301.[PMC gratis artikel]�[PubMed]
26.�Schmidt HD, Shelton RC, Duman RS. Funktionella biomarkörer för depression: diagnos, behandling och patofysiologi.�Neuropsychopharm.�2011;36(12):2375�2394.�[PMC gratis artikel]�[PubMed]
27.�J Brand S, Möller M, H Harvey B. En genomgång av biomarkörer i humör och psykotiska störningar: en dissektion av kliniska kontra prekliniska korrelat.�Curr Neuropharmacol.�2015;13(3):324�368.[PMC gratis artikel]�[PubMed]
28.�Lopresti AL, Maker GL, Hood SD, Drummond PD. En översyn av perifera biomarkörer vid allvarlig depression: potentialen hos biomarkörer för inflammatorisk och oxidativ stress.�Prog Neuropsychopharmacol Biol Psychiatry.�2014;48:102�111.�[PubMed]
29.�Fu CH, Steiner H, Costafreda SG. Prediktiva neurala biomarkörer för klinisk respons vid depression: en metaanalys av funktionella och strukturella neuroimagingstudier av farmakologiska och psykologiska terapier.�Neurobiol Dis.�2013;52:75�83.�[PubMed]
30.�Mamdani F, Berlim M, Beaulieu M, Labbe A, Merette C, Turecki G. Genuttrycksbiomarkörer för svar på citaloprambehandling vid egentlig depression.�Transl Psychiatry.�2011;1(6): e13.[PMC gratis artikel]�[PubMed]
31.�Smith RS. Makrophagteorin om depression.�Med Hypoteser.�1991;35(4):298�306.�[PubMed]
32.�Irwin MR, Miller AH. Depressiva störningar och immunitet: 20 år av framsteg och upptäckter.�Brain Behav Immun.�2007;21(4):374�383.�[PubMed]
33.�Maes M, Leonard B, Myint A, Kubera M, Verkerk R. Den nya �5-HT�-hypotesen om depression: cellmedierad immunaktivering inducerar indolamin 2,3-dioxygenas, vilket leder till lägre plasmatryptofan och en ökad syntes av skadliga tryptofankataboliter (TRYCATs), som båda bidrar till uppkomsten av depression.�Prog Neuropsychopharmacol Biol Psychiatry.�2011;35(3):702�721.[PubMed]
34.�Miller AH, Maletic V, Raison CL. Inflammation och dess missnöje: Cytokinernas roll i patofysiologin för allvarlig depression.�Biol Psykiatri.�2009;65(9):732�741.�[PMC gratis artikel]�[PubMed]
35.�Miller AH, Raison CL. Inflammationens roll i depression: från evolutionär imperativ till modern behandlingsmål.�Nat Rev Immun.�2016;16(1):22�34.�[PMC gratis artikel]�[PubMed]
36.�Raison CL, Capuron L, Miller AH. Cytokiner sjunger blues: inflammation och patogenesen av depression.�Trender Immun.�2006;27(1):24�31.�[PMC gratis artikel]�[PubMed]
37.�Raison CL, Felger JC, Miller AH. Inflammation och behandlingsresistens vid svår depression: Den perfekta stormen.�Psychiatr Times.�2013;30(9)
38.�Dowlati Y, Herrmann N, Swardfager W, et al. En metaanalys av cytokiner vid egentlig depression.�Biol Psykiatri.�2010;67(5):446�457.�[PubMed]
39.�Eyre HA, Air T, Pradhan A, et al. En metaanalys av kemokiner vid allvarlig depression.�Prog Neuropsychopharmacol Biol Psychiatry.�2016;68:1�8.�[PMC gratis artikel]�[PubMed]
40.�Haapakoski R, Mathieu J, Ebmeier KP, Alenius H, Kivim�ki M. Kumulativ metaanalys av interleukiner 6 och 1?, tumörnekrosfaktor? och C-reaktivt protein hos patienter med egentlig depression.�Brain Behav Immun.�2015;49:206�215.�[PMC gratis artikel]�[PubMed]
41.�Howren MB, Lamkin DM, Suls J. Associationer av depression med C-reaktivt protein, IL-1 och IL-6: en metaanalys.�Psychosom Med.�2009;71(2):171�186.�[PubMed]
42.�Liu Y, Ho RC-M, Mak A. Interleukin (IL)-6, tumörnekrosfaktor alfa (TNF-?) och lösliga interleukin-2-receptorer (sIL-2R) är förhöjda hos patienter med egentlig depression: en meta- analys och metaregression.�J Affect Disord.�2012;139(3):230�239.�[PubMed]
43.�Strawbridge R, Arnone D, Danese A, Papadopoulos A, Herane Vives A, Cleare AJ. Inflammation och kliniskt svar på behandling vid depression: en metaanalys.�Eur Neuropsychopharmacol.�2015;25(10):1532�1543.�[PubMed]
44.�Farooq RK, Asghar K, Kanwal S, Zulqernain A. Roll av inflammatoriska cytokiner i depression: Fokus på interleukin-1? (Recension)�Biomed Rep.�2017;6(1):15�20.�[PMC gratis artikel]�[PubMed]
45.�Cattaneo A, Ferrari C, Uher R, et al. Absoluta mätningar av makrofagmigrationshämmande faktor och interleukin-1-? mRNA-nivåer förutsäger exakt behandlingssvar hos deprimerade patienter.�Int J Neuropsychopharmacol.�2016;19(10):pyw045.�[PMC gratis artikel]�[PubMed]
46.�Baune B, Smith E, Reppermund S, et al. Inflammatoriska biomarkörer förutsäger depressiva, men inte ångestsymtom under åldrande: den blivande Sydney-studien om minne och åldrande.�Psykoneuroendokrinol.�2012;37(9):1521�1530.�[PubMed]
47.�Fornaro M, Rocchi G, Escelsior A, Contini P, Martino M. Kan olika cytokintrender hos deprimerade patienter som får duloxetin indikera differentiell biologisk bakgrund.�J Affect Disord.�2013;145(3):300�307.�[PubMed]
48.�Hernandez ME, Mendieta D, Martinez-Fong D, et al. Variationer i cirkulerande cytokinnivåer under 52 veckors behandling med SSRI för egentlig depression.�Eur Neuropsychopharmacol.�2008;18(12):917�924.�[PubMed]
49.�Hannestad J, DellaGioia N, Bloch M. Effekten av antidepressiv medicinbehandling på serumnivåer av inflammatoriska cytokiner: en metaanalys.�Neuropsykofarmakologi.�2011;36(12):2452�2459.[PMC gratis artikel]�[PubMed]
50.�Hiles SA, Attia J, Baker AL. Förändringar i interleukin-6, C-reaktivt protein och interleukin-10 hos personer med depression efter antidepressiv behandling: en metaanalys.�Brain Behav Immun; Presenteras vid: 17:e årsmötet för PsychoNeuroImmunology Research Society PsychoNeuroImmunology: Crossing Disciplines to Combat Disease; 2012. sid. S44.
51.�Harley J, Luty S, Carter J, Mulder R, Joyce P. Förhöjt C-reaktivt protein vid depression: En prediktor för bra långsiktigt resultat med antidepressiva medel och dåligt resultat med psykoterapi.�J Psychopharmacol.�2010;24(4):625�626.�[PubMed]
52.�Uher R, Tansey KE, Dew T, et al. En inflammatorisk biomarkör som en differentiell prediktor för resultatet av depressionsbehandling med escitalopram och nortriptylin.�Am J Psychiatry.�2014;171(2):1278�1286.[PubMed]
53.�Chang HH, Lee IH, Gean PW, et al. Behandlingssvar och kognitiv försämring vid egentlig depression: samband med C-reaktivt protein.�Brain Behav Immun.�2012;26(1):90�95.�[PubMed]
54.�Raison CL, Rutherford RE, Woolwine BJ, et al. En randomiserad kontrollerad studie av tumörnekrosfaktorantagonisten infliximab för behandlingsresistent depression: rollen av inflammatoriska biomarkörer vid baslinjen.�JAMA Psykiatri.�2013;70(1):31�41.�[PMC gratis artikel]�[PubMed]
55.�Krishnadas R, Cavanagh J. Depression: en inflammatorisk sjukdom?�J Neurol Neurosurg Psychiatry.�2012;83(5):495�502.�[PubMed]
56.�Raison CL, Miller AH. Är depression en inflammatorisk sjukdom?�Curr Psychiatry Rep.�2011;13(6):467�475.�[PMC gratis artikel]�[PubMed]
57.�Simon N, McNamara K, Chow C, et al. En detaljerad undersökning av cytokinavvikelser vid allvarlig depressiv sjukdom.�Eur Neuropsychopharmacol.�2008;18(3):230�233.�[PMC gratis artikel]�[PubMed]
58.�Dahl J, Ormstad H, Aass HC, et al. Plasmanivåerna av olika cytokiner ökar under pågående depression och reduceras till normala nivåer efter återhämtning.�Psykoneuroendokrinol.�2014;45:77�86.�[PubMed]
59.�Stelzhammer V, Haenisch F, Chan MK, et al. Proteomiska förändringar i serum vid första debut, patienter som inte har några antidepressiva läkemedel.�Int J Neuropsychopharmacol.�2014;17(10):1599�1608.�[PubMed]
60.�Liu Y, HO RCM, Mak A. Interleukin (IL)-17s roll i ångest och depression hos patienter med reumatoid artrit.�Int J Rheum Dis.�2012;15(2):183�187.�[PubMed]
61.�Diniz BS, Sibille E, Ding Y, et al. Plasmabiosignatur och hjärnpatologi relaterad till ihållande kognitiv funktionsnedsättning vid depression i sena livet.�Mol Psykiatri.�2015;20(5):594�601.�[PMC gratis artikel][PubMed]
62.�Janelidze S, Ventorp F, Erhardt S, et al. Förändrade kemokinnivåer i cerebrospinalvätskan och plasman hos självmordsförsökare.�Psykoneuroendokrinol.�2013;38(6):853�862.�[PubMed]
63.�Powell TR, Schalkwyk LC, Heffernan AL, et al. Tumörnekrosfaktor och dess mål i den inflammatoriska cytokinvägen identifieras som förmodade transkriptomiska biomarkörer för escitalopramsvar.�Eur Neuropsychopharmacol.�2013;23(9):1105�1114.�[PubMed]
64.�Wong M, Dong C, Maestre-Mesa J, Licinio J. Polymorfismer i inflammationsrelaterade gener är associerade med mottaglighet för allvarlig depression och antidepressiv respons.�Mol Psykiatri.�2008;13(8):800�812.�[PMC gratis artikel]�[PubMed]
65.�Kling MA, Alesci S, Csako G, et al. Ihållande låggradigt proinflammatoriskt tillstånd hos icke-medicinerade, remitterade kvinnor med egentlig depression, vilket framgår av förhöjda serumnivåer av akutfasproteinerna C-reaktivt protein och serumamyloid A.�Biol Psykiatri.�2007;62(4):309�313.�[PMC gratis artikel][PubMed]
66.�Schaefer M, Sarkar S, Schwarz M, Friebe A. Löslig intracellulär adhesionsmolekyl-1 hos patienter med unipolära eller bipolära affektiva störningar: resultat från en pilotstudie.�Neuropsykobiol.�2016;74(1):8�14.[PubMed]
67.�Dimopoulos N, Piperi C, Salonicioti A, et al. Förhöjning av plasmakoncentrationen av adhesionsmolekyler vid depression sent i livet.�Int J Geriatr Psychiatry.�2006;21(10):965�971.�[PubMed]
68.�Bocchio-Chiavetto L, Bagnardi V, Zanardini R, et al. Serum- och plasma-BDNF-nivåer vid egentlig depression: en replikationsstudie och metaanalyser.�World J Biol Psychiatry.�2010;11(6):763�773.�[PubMed]
69.�Brunoni AR, Lopes M, Fregni F. En systematisk översikt och metaanalys av kliniska studier på allvarlig depression och BDNF-nivåer: implikationer för rollen av neuroplasticitet vid depression.�Int J Neuropsychopharmacol.�2008;11(8):1169�1180.�[PubMed]
70.�Molendijk M, Spinhoven P, Polak M, Bus B, Penninx B, Elzinga B. Serum BDNF-koncentrationer som perifera manifestationer av depression: bevis från en systematisk översikt och metaanalyser på 179 föreningar.�Mol Psykiatri.�2014;19(7):791�800.�[PubMed]
71.�Sen S, Duman R, Sanacora G. Serum hjärnhärledd neurotrofisk faktor, depression och antidepressiva mediciner: metaanalyser och implikationer.�Biol Psykiatri.�2008;64(6):527�532.�[PMC gratis artikel][PubMed]
72.�Zhou L, Xiong J, Lim Y, et al. Uppreglering av blod proBDNF och dess receptorer vid allvarlig depression.�J Affect Disord.�2013;150(3):776�784.�[PubMed]
73.�Chen YW, Lin PY, Tu KY, Cheng YS, Wu CK, Tseng PT. Betydligt lägre nervtillväxtfaktornivåer hos patienter med egentlig depression än hos friska försökspersoner: en metaanalys och systematisk översikt.�Neuropsykiatri Dis Treat.�2014;11:925�933.�[PMC gratis artikel]�[PubMed]
74.�Lin PY, Tseng PT. Minskade nivåer av neurotrofisk faktor härledda från gliacellinjer hos patienter med depression: en metaanalytisk studie.�J Psychiatr Res.�2015;63:20�27.�[PubMed]
75.�Warner-Schmidt JL, Duman RS. VEGF som ett potentiellt mål för terapeutisk intervention vid depression.�Curr Op Pharmacol.�2008;8(1):14�19.�[PMC gratis artikel]�[PubMed]
76.�Carvalho AF, K�hler CA, McIntyre RS, et al. Perifer vaskulär endoteltillväxtfaktor som en ny biomarkör för depression: en metaanalys.�Psykoneuroendokrinol.�2015;62:18�26.�[PubMed]
77.�Tseng PT, Cheng YS, Chen YW, Wu CK, Lin PY. Ökade nivåer av vaskulär endotelial tillväxtfaktor hos patienter med egentlig depression: en metaanalys.�Eur Neuropsychopharmacol.�2015;25(10):1622�1630.�[PubMed]
78.�Carvalho L, Torre J, Papadopoulos A, et al. Brist på klinisk terapeutisk nytta av antidepressiva medel är associerad övergripande aktivering av det inflammatoriska systemet.�J Affect Disord.�2013;148(1):136�140.�[PubMed]
79.�Clark-Raymond A, Meresh E, Hoppensteadt D, et al. Vaskulär endoteltillväxtfaktor: Potentiell prediktor för behandlingssvar vid allvarlig depression.�World J Biol Psychiatry.�2015:1�11.�[PubMed]
80.�Isung J, Mobarrez F, Nordstr�m P, �sberg M, Jokinen J. Låg plasma vaskulär endotelial tillväxtfaktor (VEGF) associerad med fullbordat självmord.�World J Biol Psychiatry.�2012;13(6):468�473.�[PubMed]
81.�Buttensch�n HN, Foldager L, Elfving B, Poulsen PH, Uher R, Mors O. Neurotrofiska faktorer vid depression som svar på behandling.�J Affect Disord.�2015;183:287�294.�[PubMed]
82.�Szcz?sny E, ?lusarczyk J, G?ombik K, et al. Möjligt bidrag från IGF-1 till depressiv sjukdom.�Pharmacol Rep.�2013;65(6):1622�1631.�[PubMed]
83.�Tu KY, Wu MK, Chen YW, et al. Betydligt högre perifer insulinliknande tillväxtfaktor-1-nivåer hos patienter med egentlig depression eller bipolär sjukdom än hos friska kontroller: en metaanalys och granskning under Guideline of PRISMA.�Med.�2016;95(4):e2411.�[PMC gratis artikel]�[PubMed]
84.�Wu CK, Tseng PT, Chen YW, Tu KY, Lin PY. Betydligt högre perifer fibroblasttillväxtfaktor-2-nivåer hos patienter med egentlig depression: En preliminär metaanalys enligt MOOSE-riktlinjer.�Med.�2016;95(33):e4563.�[PMC gratis artikel]�[PubMed]
85.�He S, Zhang T, Hong B, et al. Minskade serumfibroblasttillväxtfaktor-2-nivåer hos patienter före och efter behandling med egentlig depression.�Neurosci Lett.�2014;579:168�172.�[PubMed]
86.�Dwivedi Y, Rizavi HS, Conley RR, Roberts RC, Tamminga CA, Pandey GN. Förändrat genuttryck av hjärnhärledd neurotrofisk faktor och receptortyrosinkinas B i hjärnan efter slakt hos självmordspersoner.�Arch Gen Psychiatry.�2003;60(8):804�815.�[PubMed]
87.�Srikanthan K, Feyh A, Visweshwar H, Shapiro JI, Sodhi K. Systematisk granskning av biomarkörer för metabolt syndrom: En panel för tidig upptäckt, hantering och riskstratifiering i befolkningen i West Virginia.�Int J Med Sci.�2016;13(1): 25.�[PMC gratis artikel]�[PubMed]
88.�Lu XY. Leptinhypotesen om depression: en potentiell koppling mellan humörstörningar och fetma?�Curr Op Pharmacol.�2007;7(6):648�652.�[PMC gratis artikel]�[PubMed]
89.�Wittekind DA, Kluge M. Ghrelin i psykiatriska störningar�En recension.�Psykoneuroendokrinol.�2015;52:176�194.�[PubMed]
90.�Kan C, Silva N, Golden SH, et al. En systematisk översikt och metaanalys av sambandet mellan depression och insulinresistens.�Diabetesvård.�2013;36(2):480�489.�[PMC gratis artikel]�[PubMed]
91.�Liu X, Li J, Zheng P, et al. Plasmalipidomik avslöjar potentiella lipidmarkörer för allvarlig depressiv sjukdom.�Anal Bioanal Chem.�2016;408(23):6497�6507.�[PubMed]
92.�Lustman PJ, Anderson RJ, Freedland KE, De Groot M, Carney RM, Clouse RE. Depression och dålig glykemisk kontroll: en metaanalytisk översikt av litteraturen.�Diabetesvård.�2000;23(7):934�942.�[PubMed]
93.�Maes M. Bevis för ett immunsvar vid allvarlig depression: en översikt och hypotes.�Prog NeuroPsychopharmacol Biol Psychiatry.�1995;19(1):11�38.�[PubMed]
94.�Zheng H, Zheng P, Zhao L, et al. Förutsägande diagnos av egentlig depression med hjälp av NMR-baserad metabolomik och minsta kvadraters stödvektormaskin.�Clinica Chimica Acta.�2017;464:223�227.[PubMed]
95.�Xia Q, Wang G, Wang H, Xie Z, Fang Y, Li Y. Studie av metabolism av glukos och lipider hos patienter med depression i första avsnittet.�J Clin Psykiatri.�2009;19:241�243.
98.�Salamone JD, Correa M, Yohn S, Cruz LL, San Miguel N, Alatorre L. Farmakologin för ansträngningsrelaterat valbeteende: Dopamin, depression och individuella skillnader.�Beteende processer.�2016;127:3�17.�[PubMed]
99.�Coplan JD, Gopinath S, Abdallah CG, Berry BR. En neurobiologisk hypotes om behandlingsresistent depression�mekanismer för selektiv serotoninåterupptagshämmare bristande effekt.�Front Behav Neurosci.�2014;8:189.�[PMC gratis artikel]�[PubMed]
100.�Popa D, Cerdan J, Rep�rant C, et al. En longitudinell studie av 5-HT-utflöde under kronisk fluoxetinbehandling med en ny teknik för kronisk mikrodialys i en mycket känslomässig musstam.�Eur J Pharmacol.�2010;628(1):83�90.�[PubMed]
101.�Atake K, Yoshimura R, Hori H, et al. Duloxetin, en selektiv noradrenalinåterupptagshämmare, ökade plasmanivåerna av 3-metoxi-4-hydroxifenylglykol men inte homovanillinsyra hos patienter med egentlig depression.�Clin Psychopharmacol Neurosci.�2014;12(1):37�40.�[PMC gratis artikel][PubMed]
102.�Ueda N, Yoshimura R, Shinkai K, Nakamura J. Plasmanivåer av katekolaminmetaboliter förutsäger svaret på sulpirid eller fluvoxamin vid allvarlig depression.�Farmakopsykiatri.�2002;35(05):175�181.[PubMed]
103.�Yamana M, Atake K, Katsuki A, Hori H, Yoshimura R. Blodbiologiska markörer för förutsägelse av escitalopramsvar hos patienter med egentlig depression: preliminär studie.�J Depressiv ångest.�2016;5: 222.
104.�Parker KJ, Schatzberg AF, Lyons DM. Neuroendokrina aspekter av hyperkortisolism vid allvarlig depression.�Horm Behav.�2003;43(1):60�66.�[PubMed]
105.�Stetler C, Miller GE. Depression och hypotalamus-hypofys-binjureaktivering: en kvantitativ sammanfattning av fyra decennier av forskning.�Psychosom Med.�2011;73(2):114�126.�[PubMed]
106.�Herane Vives A, De Angel V, Papadopoulos A, et al. Relationen mellan kortisol, stress och psykiatrisk sjukdom: Nya insikter med hjälp av håranalys.�J Psychiatr Res.�2015;70:38�49.�[PubMed]
107.�Fischer S, Strawbridge R, Vives AH, Cleare AJ. Kortisol som en prediktor för psykologisk terapisvar vid depressiva sjukdomar: systematisk översikt och metaanalys.�Br J Psykiatri.�2017;210(2):105�109.�[PubMed]
108.�Anacker C, Zunszain PA, Carvalho LA, Pariante CM. Glukokortikoidreceptorn: nyckeln till depression och antidepressiv behandling?�Psykoneuroendokrinologi.�2011;36(3):415�425.�[PMC gratis artikel][PubMed]
109.�Markopoulou K, Papadopoulos A, Juruena MF, Poon L, Pariante CM, Cleare AJ. Förhållandet mellan kortisol/DHEA vid behandlingsresistent depression.�Psykoneuroendokrinol.�2009;34(1):19�26.�[PubMed]
110.�Joffe RT, Pearce EN, Hennessey JV, Ryan JJ, Stern RA. Subklinisk hypotyreos, humör och kognition hos äldre vuxna: en recension.�Int J Geriatr Psychiatry.�2013;28(2):111�118.�[PMC gratis artikel][PubMed]
111.�Duval F, Mokrani MC, Erb A, et al. Kronobiologisk hypotalamus�hypofys�status för sköldkörtelaxeln och antidepressivt resultat vid allvarlig depression.�Psykoneuroendokrinol.�2015;59:71�80.�[PubMed]
112.�Marsden W. Synaptisk plasticitet vid depression: molekylära, cellulära och funktionella korrelat.�Prog Neuropsychopharmacol Biol Psychiatry.�2013;43:168�184.�[PubMed]
113.�Duman RS, Voleti B. Signalvägar som ligger bakom patofysiologin och behandlingen av depression: nya mekanismer för snabbverkande medel.�Trender Neurosci.�2012;35(1):47�56.[PMC gratis artikel]�[PubMed]
114.�Ripke S, Wray NR, Lewis CM, et al. En megaanalys av genomomfattande associationsstudier för allvarlig depressiv sjukdom.�Mol Psykiatri.�2013;18(4):497�511.�[PMC gratis artikel]�[PubMed]
115.�Mullins N, Power R, Fisher H, et al. Polygena interaktioner med miljömässiga motgångar i etiologin för allvarlig depressiv sjukdom.�Psychol Med.�2016;46(04):759�770.�[PMC gratis artikel]�[PubMed]
116.�Lewis S. Neurologiska störningar: telomerer och depression.�Nat Rev Neurosci.�2014;15(10): 632.[PubMed]
117.�Lindqvist D, Epel ES, Mellon SH, et al. Psykiatriska störningar och leukocyttelomerlängd: underliggande mekanismer som kopplar samman psykisk sjukdom med cellulärt åldrande.�Neurosci Biobehav Rev.�2015;55:333�364.�[PMC gratis artikel]�[PubMed]
118.�McCall WV. En biomarkör för viloaktivitet för att förutsäga svar på SSRI vid egentlig depression.�J Psychiatr Res.�2015;64:19�22.�[PMC gratis artikel]�[PubMed]
119.�Schuch FB, Deslandes AC, Stubbs B, Gosmann NP, da Silva CTB, de Almeida Fleck MP. Neurobiologiska effekter av träning på allvarlig depressiv sjukdom: en systematisk översikt.�Neurosci Biobehav Rev.�2016;61:1�11.�[PubMed]
120.�Foster JA, Neufeld K-AM. Tarmhjärnans axel: hur mikrobiomet påverkar ångest och depression.�Trender Neurosci.�2013;36(5):305�312.�[PubMed]
121.�Quattrocki E, Baird A, Yurgelun-Todd D. Biologiska aspekter av sambandet mellan rökning och depression.�Harv Rev Psychiatry.�2000;8(3):99�110.�[PubMed]
122.�Maes M, Kubera M, Obuchowiczwa E, Goehler L, Brzeszcz J. Depressions multipla komorbiditeter förklaras av (neuro)inflammatoriska och oxidativa och nitrosativa stressvägar.�Neuro Endocrinol Lett.�2011;32(1):7�24.�[PubMed]
123.�Miller G, Rohleder N, Cole SW. Kronisk interpersonell stress förutsäger aktivering av pro- och antiinflammatoriska signalvägar sex månader senare.�Psychosom Med.�2009;71(1): 57.�[PMC gratis artikel][PubMed]
124.�Steptoe A, Hamer M, Chida Y. Effekterna av akut psykologisk stress på cirkulerande inflammatoriska faktorer hos människor: en översikt och metaanalys.�Brain Behav Immun.�2007;21(7):901�912.�[PubMed]
125.�Danese A, Moffitt TE, Harrington H, et al. Negativa barndomsupplevelser och vuxen riskfaktorer för åldersrelaterad sjukdom: depression, inflammation och klustring av metabola riskmarkörer.�Arch Pediatr Adolesc Med.�2009;163(12):1135�1143.�[PMC gratis artikel]�[PubMed]
126.�Danese A, Pariante CM, Caspi A, Taylor A, Poulton R. Misshandel hos barn förutsäger inflammation hos vuxna i en livsloppsstudie.�Proc Natl Acad Sci US A.�2007;104(4):1319�1324.�[PMC gratis artikel][PubMed]
127.�Danska A, Caspi A, Williams B, et al. Biologisk inbäddning av stress genom inflammationsprocesser i barndomen.�Mol Psykiatri.�2011;16(3):244�246.�[PMC gratis artikel]�[PubMed]
128.�Suzuki A, Poon L, Kumari V, Cleare AJ. Rädsla fördomar i emotionell ansiktsbehandling efter barndomstrauma som en markör för motståndskraft och sårbarhet för depression.�Barnmisshandel.�2015;20(4):240�250.�[PubMed]
129.�Strawbridge R, Young AH. HPA-axeln och kognitiv dysreglering vid humörstörningar. I: McIntyre RS, Cha DS, redaktörer.�Kognitiv funktionsnedsättning vid allvarlig depressiv sjukdom: klinisk relevans, biologiska underlag och behandlingsmöjligheter.�Cambridge: Cambridge University Press; 2016. s. 179�193.
130.�Keller J, Gomez R, Williams G, et al. HPA-axeln vid egentlig depression: kortisol, klinisk symptomatologi och genetisk variation förutsäger kognition.�Mol Psykiatri.�2016 augusti 16;�Epub.�[PMC gratis artikel]�[PubMed]
131.�Hanson ND, Owens MJ, Nemeroff CB. Depression, antidepressiva medel och neurogenes: en kritisk omvärdering.�Neuropsykofarmakol.�2011;36(13):2589�2602.�[PMC gratis artikel]�[PubMed]
132.�Chen Y, Baram TZ. Mot att förstå hur stress i tidigt liv omprogrammerar kognitiva och emotionella hjärnnätverk.�Neuropsykofarmakol.�2015;41(1):197�206.�[PMC gratis artikel]�[PubMed]
133.�Porter RJ, Gallagher P, Thompson JM, Young AH. Neurokognitiv funktionsnedsättning hos drogfria patienter med egentlig depression.�Br J Psykiatri.�2003;182:214�220.�[PubMed]
134.�Gallagher P, Robinson L, Gray J, Young A, Porter R. Neurokognitiv funktion efter remission vid egentlig depression: potentiell objektiv markör för svar?�Aust NZJ Psychiatry.�2007;41(1):54�61.�[PubMed]
135.�Pittenger C, Duman RS. Stress, depression och neuroplasticitet: en konvergens av mekanismer.�Neuropsykofarmakol.�2008;33(1):88�109.�[PubMed]
136.�Beckman L, Nyberg L, Lindenberger U, Li SC, Farde L. Den korrelativa triaden mellan åldrande, dopamin och kognition: nuvarande status och framtidsutsikter.�Neurosci Biobehav Rev.�2006;30(6):791�807.�[PubMed]
137.�Allison DJ, Ditor DS. Den vanliga inflammatoriska etiologin för depression och kognitiv funktionsnedsättning: ett terapeutiskt mål.�J Neuroinflammation.�2014;11:151.�[PMC gratis artikel]�[PubMed]
138.�Rosenblat JD, Brietzke E, Mansur RB, Maruschak NA, Lee Y, McIntyre RS. Inflammation som ett neurobiologiskt substrat för kognitiv funktionsnedsättning vid bipolär sjukdom: bevis, patofysiologi och behandlingsimplikationer.�J Affect Disord.�2015;188:149�159.�[PubMed]
139.�Krogh J, Benros ME, J�rgensen MB, Vesterager L, Elfving B, Nordentoft M. Sambandet mellan depressiva symtom, kognitiv funktion och inflammation vid allvarlig depression.�Brain Behav Immun.�2014;35:70�76.�[PubMed]
140.�Soares CN, Zitek B. Reproduktiv hormonkänslighet och risk för depression över den kvinnliga livscykeln: ett kontinuum av sårbarhet?�J Psychiatry Neurosci.�2008;33(4): 331.�[PMC gratis artikel]�[PubMed]
141.�Hiles SA, Baker AL, de Malmanche T, Attia J. En metaanalys av skillnader i IL-6 och IL-10 mellan personer med och utan depression: utforska orsakerna till heterogenitet.�Brain Behav Immun.�2012;26(7):1180�1188.�[PubMed]
142.�Fontana L, Eagon JC, Trujillo ME, Scherer PE, Klein S. Utsöndring av visceralt fett adipokine är associerat med systemisk inflammation hos överviktiga människor.�Diabetes.�2007;56(4):1010�1013.�[PubMed]
143.�Divani AA, Luo X, Datta YH, Flaherty JD, Panoskaltsis-Mortari A. Effekt av orala och vaginala hormonella preventivmedel på inflammatoriska blodbiomarkörer.�Mediators Inflamm.�2015;2015: 379501.[PMC gratis artikel]�[PubMed]
144.�Ramsey JM, Cooper JD, Penninx BW, Bahn S. Variation i serumbiomarkörer med kön och kvinnlig hormonstatus: konsekvenser för kliniska tester.�Sci Rep.�2016;6:26947.�[PMC gratis artikel]�[PubMed]
145.�Eyre H, Lavretsky H, Kartika J, Qassim A, Baune B. Modulerande effekter av antidepressiva klasser på det medfödda och adaptiva immunsystemet vid depression.�Farmakopsykiatri.�2016;49(3):85�96.[PMC gratis artikel]�[PubMed]
146.�Hiles SA, Baker AL, de Malmanche T, Attia J. Interleukin-6, C-reactive protein and interleukin-10 efter antidepressiv behandling hos personer med depression: en metaanalys.�Psychol Med.�2012;42(10):2015�2026.�[PubMed]
147.�Janssen DG, Caniato RN, Verster JC, Baune BT. En psykoneuroimmunologisk granskning av cytokiner involverade i behandlingssvaret mot antidepressiva.�Hum Psychopharmacol.�2010;25(3):201�215.�[PubMed]
148.�Artigas F. Serotoninreceptorer involverade i antidepressiva effekter.�Pharmacol Ther.�2013;137(1):119�131.�[PubMed]
149.�Lee BH, Kim YK. BDNF:s roller i patofysiologin för allvarlig depression och i antidepressiv behandling.�Psykiatriutredning.�2010;7(4):231�235.�[PMC gratis artikel]�[PubMed]
150.�Hashimoto K. Inflammatoriska biomarkörer som differentiella prediktorer för antidepressivt svar.�Int J Mol Sci.�2015;16(4):7796�7801.�[PMC gratis artikel]�[PubMed]
151.�Goldberg D. The heterogenity of �major depression��Världspsykiatri.�2011;10(3):226�228.[PMC gratis artikel]�[PubMed]
152.�Arnow BA, Blasey C, Williams LM, et al. Depressionsundertyper för att förutsäga antidepressivt svar: en rapport från iSPOT-D-studien.�Am J Psychiatry.�2015;172(8):743�750.�[PubMed]
153.�Kunugi H, Hori H, Ogawa S. Biokemiska markörer som subtypar allvarlig depressiv sjukdom.�Psykiatri Clin Neurosci.�2015;69(10):597�608.�[PubMed]
154.�Baune B, Stuart M, Gilmour A, et al. Förhållandet mellan undertyper av depression och hjärt-kärlsjukdom: en systematisk genomgång av biologiska modeller.�Transl Psychiatry.�2012;2(3): e92.[PMC gratis artikel]�[PubMed]
155.�Vogelzangs N, Duivis HE, Beekman AT, et al. Samband mellan depressiva sjukdomar, depressionsegenskaper och antidepressiv medicin med inflammation.�Transl Psychiatry.�2012;2: e79.[PMC gratis artikel]�[PubMed]
156.�Lamers F, Vogelzangs N, Merikangas K, De Jonge P, Beekman A, Penninx B. Bevis för en differentiell roll av HPA-axelfunktion, inflammation och metabolt syndrom i melankolisk kontra atypisk depression.�Mol Psykiatri.�2013;18(6):692�699.�[PubMed]
157.�Penninx BW, Milaneschi Y, Lamers F, Vogelzangs N. Förstå de somatiska konsekvenserna av depression: biologiska mekanismer och rollen av depressionssymptomprofil.�BMC Med.�2013;11(1): 1.[PMC gratis artikel]�[PubMed]
158.�Capuron L, Su S, Miller AH, et al. Depressiva symtom och metabolt syndrom: Är inflammation den underliggande länken?�Biol Psykiatri.�2008;64(10):896�900.�[PMC gratis artikel]�[PubMed]
159.�Dantzer R, O'Connor JC, Freund GG, Johnson RW, Kelley KW. Från inflammation till sjukdom och depression: när immunsystemet lägger ner hjärnan.�Nat Rev Neurosci.�2008;9(1):46�56.[PMC gratis artikel]�[PubMed]
160.�Maes M, Berk M, Goehler L, et al. Depression och sjukdomsbeteende är Janus-ansikte svar på delade inflammatoriska vägar.�BMC Med.�2012;10:66.�[PMC gratis artikel]�[PubMed]
161.�Merikangas KR, Jin R, He JP, et al. Prevalens och korrelat av bipolär spektrumstörning i World Mental Health Survey Initiative.�Arch Gen Psychiatry.�2011;68(3):241�251.�[PMC gratis artikel][PubMed]
162.�Hirschfeld RM, Lewis L, Vornik LA. Uppfattningar och effekter av bipolär sjukdom: hur långt har vi egentligen kommit? Resultat från National depressive and mano-depressive association 2000 undersökning av individer med bipolär sjukdom.�J Clin Psykiatri.�2003;64(2):161�174.�[PubMed]
163.�Young AH, MacPherson H. Detektion av bipolär sjukdom.�Br J Psykiatri.�2011;199(1):3�4.[PubMed]
164.�Vhringer PA, Perlis RH. Diskriminering mellan bipolär sjukdom och egentlig depression.�Psykiatriker Clin North Am.�2016;39(1):1�10.�[PubMed]
165.�Becking K, Spijker AT, Hoencamp E, Penninx BW, Schoevers RA, Boschloo L. Störningar i hypotalamus-hypofys-binjureaxeln och immunologisk aktivitet som skiljer mellan unipolära och bipolära depressiva episoder.�PLoS One.�2015;10(7):e0133898.�[PMC gratis artikel]�[PubMed]
166.�Huang TL, Lin FC. Högkänsliga C-reaktiva proteinnivåer hos patienter med egentlig depression och bipolär mani.�Prog NeuroPsychopharmacol Biol Psychiatry.�2007;31(2):370�372.�[PubMed]
167.�Angst J, Gamma A, Endrass J. Riskfaktorer för de bipolära och depressionsspektra.�Acta Psychiatr Scand.�2003;418:15�19.�[PubMed]
168.�Fekadu A, Wooderson S, Donaldson C, et al. Ett multidimensionellt verktyg för att kvantifiera behandlingsmotstånd vid depression: Maudsleys iscensättningsmetod.�J Clin Psykiatri.�2009;70(2): 177.�[PubMed]
169.�Papakostas G, Shelton R, Kinrys G, et al. Bedömning av ett multi-assay, serumbaserat biologiskt diagnostiskt test för egentlig depression: en pilot- och replikationsstudie.�Mol Psykiatri.�2013;18(3):332�339.�[PubMed]
170.�Fan J, Han F, Liu H. Utmaningar med stordataanalys.�Natl Sci Rev.�2014;1(2):293�314.[PMC gratis artikel]�[PubMed]
171.�Li L, Jiang H, Qiu Y, Ching WK, Vassiliadis VS. Upptäckt av metabolitbiomarkörer: flödesanalys och reaktions-reaktionsnätverk.�BMC Syst Biol.�2013;7(Suppl 2):S13.�[PMC gratis artikel][PubMed]
172.�Patel MJ, Khalaf A, Aizenstein HJ. Att studera depression med hjälp av bild- och maskininlärningsmetoder.�NeuroImage Clin.�2016;10:115�123.�[PMC gratis artikel]�[PubMed]
173.�Lanquillon S, Krieg JC, Bening-Abu-Shach U, Vedder H. Cytokinproduktion och behandlingssvar vid allvarlig depressiv sjukdom.�Neuropsykofarmakol.�2000;22(4):370�379.�[PubMed]
174.�Lindqvist D, Janelidze S, Erhardt S, Tr�skman-Bendz L, Engstr�m G, Brundin L. CSF-biomarkörer i självmordsförsökare�a principal component analysis.�Acta Psychiatr Scand.�2011;124(1):52�61.�[PubMed]
175.�Hidalgo-Mazzei D, Murru A, Reinares M, Vieta E, Colom F. Big data in mental health: a challenging fragmented future.�Världspsykiatri.�2016;15(2):186�187.�[PMC gratis artikel]�[PubMed]
176.�Konsortium C-DGotPG Identifiering av risklägen med delade effekter på fem stora psykiatriska störningar: en genomomfattande analys.�Lancet.�2013;381(9875):1371�1379.�[PMC gratis artikel]�[PubMed]
177.�Dipnall JF, Pasco JA, Berk M, et al. Kombinerar datautvinning, maskininlärning och traditionell statistik för att upptäcka biomarkörer associerade med depression.�PLoS One.�2016;11(2):e0148195.�[PMC gratis artikel][PubMed]
178.�K�hler O, Benros ME, Nordentoft M, et al. Effekt av antiinflammatorisk behandling på depression, depressiva symtom och biverkningar: en systematisk översikt och metaanalys av randomiserade kliniska prövningar.�JAMA Psykiatri.�2014;71(12):1381�1391.�[PubMed]
179.�Wolkowitz OM, Reus VI, Chan T, et al. Antiglukokortikoidbehandling av depression: dubbelblind ketokonazol.�Biol Psykiatri.�1999;45(8):1070�1074.�[PubMed]
180.�McAllister-Williams RH, Anderson IM, Finkelmeyer A, et al. Antidepressiv förstärkning med metyrapon för behandlingsresistent depression (ADD-studien): en dubbelblind, randomiserad, placebokontrollerad studie.�Lancet Psychiatry.�2016;3(2):117�127.�[PubMed]
181.�Gallagher P, Young AH. Mifepriston (RU-486) behandling för depression och psykos: En genomgång av de terapeutiska implikationerna.�Neuropsykiatri Dis Treat.�2006;2(1):33�42.�[PMC gratis artikel]�[PubMed]
182.�Otte C, Hinkelmann K, Moritz S, et al. Modulering av mineralokortikoidreceptorn som tilläggsbehandling vid depression: en randomiserad, dubbelblind, placebokontrollerad proof-of-concept-studie.�J Psychiatr Res.�2010;44(6):339�346.�[PubMed]
183.�Ozbolt LB, Nemeroff CB. HPA-axelmodulering vid behandling av humörstörningar.�Psykiatrisk sjukdom.�2013;51:1147�1154.
184.�Walker AK, Budac DP, Bisulco S, et al. NMDA-receptorblockad av ketamin upphäver lipopolysackarid-inducerat depressivt beteende hos C57BL/6J-möss.�Neuropsykofarmakol.�2013;38(9):1609�1616.�[PMC gratis artikel]�[PubMed]
185.�Lesp�rance F, Frasure-Smith N, St-Andr� E, Turecki G, Lesp�rance P, Wisniewski SR. Effekten av omega-3-tillskott för allvarlig depression: en randomiserad kontrollerad studie.�J Clin Psykiatri.�2010;72(8):1054�1062.�[PubMed]
186.�Kim S, Bae K, Kim J, et al. Användning av statiner för behandling av depression hos patienter med akut koronarsyndrom.�Transl Psychiatry.�2015;5(8):e620.�[PMC gratis artikel]�[PubMed]
187.�Shishehbor MH, Brennan ML, Aviles RJ, et al. Statiner främjar potenta systemiska antioxidanteffekter genom specifika inflammatoriska vägar.�Cirkulation.�2003;108(4):426�431.�[PubMed]
188.�Mercier A, Auger-Aubin I, Lebeau JP, et al. Bevis på förskrivning av antidepressiva läkemedel för icke-psykiatriska tillstånd i primärvården: en analys av riktlinjer och systematiska översikter.�BMC Family Practice.�2013;14(1): 55.�[PMC gratis artikel]�[PubMed]
189.�Freland L, Beaulieu JM. Hämning av GSK3 av litium, från enstaka molekyler till signalnätverk.�Främre Mol Neurosci.�2012;5:14.�[PMC gratis artikel]�[PubMed]
190.�Horowitz MA, Zunszain PA. Neuroimmuna och neuroendokrina abnormiteter vid depression: två sidor av samma mynt.�Ann NY Acad Sci.�2015;1351(1):68�79.�[PubMed]
191.�Juruena MF, Cleare AJ. Överlappning mellan atypisk depression, säsongsbetonad affektiv störning och kroniskt trötthetssyndrom.�Rev Bras Psiquiatr.�2007;29:S19�S26.�[PubMed]
192.�Castr�n E, Kojima M. Hjärnhärledd neurotrofisk faktor vid humörstörningar och antidepressiva behandlingar.�Neurobiol Dis.�2017;97(Pt B):119�126.�[PubMed]
193.�Pan A, Keum N, Okereke OI, et al. Dubbelriktad association mellan depression och metabolt syndrom en systematisk översikt och metaanalys av epidemiologiska studier.�Diabetesvård.�2012;35(5):1171�1180.�[PMC gratis artikel]�[PubMed]
194.�Carvalho AF, Rocha DQ, McIntyre RS, et al. Adipokines som nya biomarkörer för depression: en systematisk översikt och metaanalys.�J Psychiatric Res.�2014;59:28�37.�[PubMed]
195.�Wise T, Cleare AJ, Herane A, Young AH, Arnone D. Diagnostisk och terapeutisk användbarhet av neuroimaging vid depression: en översikt.�Neuropsykiatri Dis Treat.�2014;10:1509�1522.[PMC gratis artikel]�[PubMed]
196.�Tamatam A, Khanum F, Bawa AS. Genetiska biomarkörer för depression.�Indiska J Hum Genet.�2012;18(1): 20.�[PMC gratis artikel]�[PubMed]
197.�Yoshimura R, Nakamura J, Shinkai K, Ueda N. Kliniskt svar på antidepressiv behandling och 3-metoxi-4-hydroxifenylglykolnivåer: minirecension.�Prog Neuropsychopharmacol Biol Psychiatry.�2004;28(4):611�616.�[PubMed]
198.�Pierscionek T, Adekunte O, Watson S, Ferrier N, Alabi A. Kortikosteroidernas roll i det antidepressiva svaret.�ChronoPhys Ther.�2014;4:87�98.
199.�Hage MP, Azar ST. Kopplingen mellan sköldkörtelfunktion och depression.�J Thyroid Res.�2012;2012:590648.�[PMC gratis artikel]�[PubMed]
200.�Dunn EC, Brown RC, Dai Y, et al. Genetiska bestämningsfaktorer för depression: senaste fynd och framtida riktningar.�Harv Rev Psychiatry.�2015;23(1): 1.�[PMC gratis artikel]�[PubMed]
201.�Yang CC, Hsu YL. En recension av accelerometribaserade bärbara rörelsedetektorer för övervakning av fysisk aktivitet.�Sensorer.�2010;10(8):7772�7788.�[PMC gratis artikel]�[PubMed]
Artrit smärta är ett komplext fenomen som involverar intrikat neurofysiologisk bearbetning på alla nivåer av smärtvägen. De tillgängliga behandlingsalternativen för att lindra ledvärk är ganska begränsade, och de flesta artritpatienter rapporterar endast blygsam smärtlindring med nuvarande behandlingar. En bättre förståelse av de neurala mekanismer som är ansvariga för muskel- och skelettsmärta och identifiera nya mål kommer att hjälpa till att utveckla framtida farmakologiska terapier. Den här artikeln går igenom en del av den senaste forskningen om faktorer som bidrar till ledvärk och täcker områden som cannabinoider, proteinasaktiverade receptorer, natriumkanaler, cytokiner och transienta receptorpotentialkanaler. Den framväxande hypotesen att artros kan ha en neuropatisk komponent diskuteras också.
Beskrivning
Världshälsoorganisationen rankar muskuloskeletala störningar som den vanligaste orsaken till funktionshinder i den moderna världen, vilket påverkar en av tre vuxna [1]. Ännu mer alarmerande är att förekomsten av dessa sjukdomar ökar medan vår kunskap om deras bakomliggande orsaker är ganska rudimentär.
Fig. 1 En schematisk bild som illustrerar några av målen som är kända för att modulera ledsmärta. Neuromodulatorer kan frigöras från nervterminaler såväl som mastceller och makrofager för att förändra afferent mekanokänslighet. Endovanilloider, syra och skadlig värme kan aktivera transienta receptorpotentiala vanilloid typ 1 (TRPV1) jonkanaler vilket leder till frisättning av algogen substans P (SP), som därefter binder till neurokinin-1 (NK1) receptorer. Proteaser kan klyva och stimulera proteasaktiverade receptorer (PAR). Hittills har PAR2 och PAR4 visat sig sensibilisera gemensamma primära afferenter. Endocannabinoiden anandamid (AE) produceras på begäran och syntetiseras från N-arakidonoylfosfatidyletanolamin (NAPE) under den enzymatiska effekten av fosfolipaser. En del av AE binder sedan till cannabinoid-1 (CB1) receptorer vilket leder till neuronal desensibilisering. Obundet AE tas snabbt upp av en anandamidmembrantransportör (AMT) innan den bryts ned av ett fettsyraamidhydrolas (FAAH) till etanolamin (Et) och arakidonsyra (AA). Cytokinerna tumörnekrosfaktor-?(TNF-?), interleukin-6 (IL-6) och interleukin1-beta (IL-1?) kan binda till sina respektive receptorer för att öka smärtöverföringen. Slutligen är tetrodotoxin (TTX)-resistenta natriumkanaler (Nav1.8) involverade i neuronal sensibilisering.
Patienter längtar efter deras kronisk smärta att försvinna; men för närvarande ordinerade smärtstillande medel är i stort sett ineffektiva och åtföljs av ett brett spektrum av oönskade biverkningar. Som sådan lider miljontals människor världen över av de försvagande effekterna av ledvärk, för vilka det inte finns någon tillfredsställande behandling [2].
Mer än 100 olika former av artrit har artros (OA) som den vanligaste. OA är en progressivt degenerativ ledsjukdom som orsakar kronisk smärta och funktionsförlust. Vanligtvis är OA ledens oförmåga att reparera skador effektivt som svar på överdrivna krafter som utsätts på den. De biologiska och psykosociala faktorerna som innefattar kronisk OA-smärta är inte välkända, även om pågående forskning avslöjar sjukdomssymtomens komplexa natur [2]. Aktuella terapier, såsom icke-steroida antiinflammatoriska läkemedel (NSAID), ger viss symptomatisk lindring, minskar smärtan under korta perioder, men lindrar inte smärta under patientens livslängd. Dessutom kan höga doser NSAID inte tas upprepade gånger under många år, eftersom detta kan leda till njurtoxicitet och gastrointestinala blödningar.
Traditionellt har artritforskningen till stor del fokuserat på ledbrosket som ett primärt mål för den terapeutiska utvecklingen av nya OA-läkemedel för sjukdomsmodifiering. Detta kondrogena fokus har kastat nytt ljus över de komplicerade biokemiska och biomekaniska faktorerna som påverkar kondrocytbeteende i sjuka leder. Men eftersom ledbrosket är aneuralt och avaskulärt är det osannolikt att denna vävnad är källan till OA-smärta. Detta faktum, tillsammans med fynden att det inte finns någon korrelation mellan skadan av ledbrosk och smärta hos OA-patienter [3,4] eller prekliniska modeller av OA [5], har orsakat en förändring i fokus för att utveckla läkemedel för effektiv smärtkontroll . Den här artikeln kommer att granska de senaste rönen inom ledsmärtaforskning och lyfta fram några av de framväxande målen som kan vara framtiden för smärtbehandling av artrit (sammanfattat i fig. 1)
cytokiner
Verkningarna av olika cytokiner i gemensamma neurofysiologiska studier har varit ganska framträdande nyligen. Interleukin-6 (IL-6), till exempel, är ett cytokin som typiskt binder till den membranbundna IL-6-receptorn (IL-6R). IL-6 kan också signalera genom att binda till ett lösligt IL-6R (SIL-6R) för att producera ett IL-6/sIL-6R-komplex. Detta IL-6/sIL-6R-komplex binds därefter till en transmembranglykoproteinsubenhet 130(gp130), och tillåter därigenom IL-6 att signalera i celler som inte konstitutivt uttrycker membranbundet IL-6R [25,26]. IL-6 och SIL-6R är nyckelspelare vid systemisk inflammation och artrit, eftersom uppreglering av båda har hittats i RA-patienters serum och ledvätska [27,29]. Nyligen observerade Vazquez et al. att samtidig administrering av IL-6/sIL-6R i råttknän orsakade inflammationsframkallad smärta, vilket avslöjades av en ökning av svaret från spinal dorsalhornsneuroner på mekanisk stimulering av knät och andra delar av bakbenet [30]. Spinal neuron hyperexcitabilitet sågs också när IL-6/sIL-6R applicerades lokalt på ryggmärgen. Spinal applicering av lösligt gp130 (som skulle torka upp IL-6/sIL-6R-komplex och därigenom minska trans-signalering) hämmade IL-6/sIL-6R-inducerad central sensibilisering. Akut applicering av enbart lösligt gpl130 reducerade emellertid inte de neuronala svaren på redan etablerad ledinflammation.
De transienta receptorpotentialkanalerna (TRP) är icke-selektiva katjonkanaler som fungerar som integratorer av olika fysiologiska och patofysiologiska processer. Förutom värmesensation, kemosensation och mekanosensation är TRP-kanaler involverade i moduleringen av smärta och inflammation. Till exempel har TRP vanilloid-1 (TRPV1) jonkanaler visat sig bidra till ledinflammatorisk smärta eftersom termisk hyperalgesi inte var framkallande i TRPV1 mono artritiska möss [31]. På liknande sätt är TRP ankyrin-1 (TRPA1) jonkanaler involverade i artritisk mekanöverkänslighet eftersom blockad av receptorn med selektiva antagonister dämpade mekanisk smärta i Freunds kompletta adjuvansmodellinflammation [32,33]. Ytterligare bevis på att TRPV1 kan vara involverad i neurotransmissionen av OA-smärta kommer från studier där neuronalt TRPV1-uttryck är förhöjt i natriummonojodacetatmodellen av OA [34]. Dessutom reducerade systemisk administrering av TRPV1-antagonisten A-889425 den framkallade och spontana aktiviteten hos spinal-wide dynamiska intervall och nociception-specifika neuroner i monojodacetatmodellen [35]. Dessa data tyder på att endovanilloider kan vara involverade i centrala sensibiliseringsprocesser associerade med OA-smärta.
Det är för närvarande kända för att finnas minst fyra polymorfismer i genen som kodar för TRPV1, vilket leder till en förändring i strukturen av jonkanalen och försämrad funktion. En speciell polymorfism (rs8065080) förändrar känsligheten hos TRPV1 för capsaicin, och individer som bär denna polymorfism är mindre känsliga för termisk hyperalgesi [36]. En nyligen genomförd studie undersökte om OA-patienter med rs8065080 polymorfism upplevde förändrad smärtuppfattning baserat på denna genetiska anomali. Forskargruppen fann att patienter med asymtomatisk knä-OA var mer benägna att bära på rs8065080-genen än patienter med smärtsamma leder [37]. Denna observation indikerar att OA-patienter med normal funktion; TRPV1-kanaler har en ökad risk för ledvärk och bekräftar åter den potentiella involveringen av TRPV1 i OA-smärta.
Slutsats
Även om hindret för att behandla artritsmärta effektivt kvarstår, görs stora språng i vår förståelse av de neurofysiologiska processer som är ansvariga för genereringen av ledvärk. Nya mål upptäcks kontinuerligt, samtidigt som mekanismerna bakom kända vägar definieras och förfinas ytterligare. Att rikta in sig på en specifik receptor eller jonkanal är sannolikt inte lösningen för att normalisera ledsmärta, utan snarare indikeras ett polyfarmacetiskt tillvägagångssätt där olika mediatorer används i kombination under specifika faser av sjukdomen. Att reda ut de funktionella kretsarna på varje nivå av smärtbanan kommer också att förbättra vår kunskap om hur ledvärk genereras. Till exempel kommer identifiering av de perifera mediatorerna av ledsmärta att tillåta oss att kontrollera nociception i leden och sannolikt undvika de centrala biverkningarna av systemiskt administrerade farmakoterapeutika.
FACETOGEN PAIN
FACETSYNDROM OCH FACETOGENISKT SMÄRT
Facetsyndrom är en artikulär störning relaterad till ländryggsfasettlederna och deras innerveringar och ger både lokal och utstrålande facetogen smärta.
Överdriven rotation, förlängning eller böjning av ryggraden (upprepad överanvändning) kan resultera i degenerativa förändringar i ledens brosk. Dessutom kan det innebära degenerativa förändringar i andra strukturer, inklusive mellankotskivan.
Smärta med och / eller begränsning av förlängning och rotation
Tenderness vid palpation
Utstrålande facetogena smärtor lokalt eller i axlarna eller övre ryggen och sällan strålar fram eller ner i armen eller i fingrarna som en hernierad skiva kan.
LUMBAR FACET SYNDROM & FACETOGENIC SMÄRT
Smärta eller ömhet i nedre delen av ryggen.
Lokal ömhet / stelhet vid sidan av ryggraden i nedre delen av ryggen.
Smärta, stelhet eller svårigheter med vissa rörelser (som att stå upprätt eller resa sig från en stol.
Smärta vid hyperextension
Refererad smärta från övre ländryggsfasettleder kan sträcka sig in i flanken, höften och övre laterala låret.
Refererad smärta från nedre ländryggsfasettleder kan penetrera djupt in i låret, i sidled och/eller baktill.
L4-L5 och L5-S1 facettleder kan hänvisa till smärta som sträcker sig in i det distala laterala benet och i sällsynta fall till foten
EVIDENSBASERAD MEDICIN
Bevisbaserad interventionsmedicin enligt kliniska diagnoser
12. Smärta som härrör från ländryggfoglederna
Abstrakt
Även om förekomsten av ett facettsyndrom länge hade ifrågasatts, är det nu allmänt accepterat som en klinisk enhet. Beroende på de diagnostiska kriterierna står de zygapofysiska lederna för mellan 5 % och 15 % av fallen av kronisk, axiell ländryggssmärta. Vanligast är facetogen smärta ett resultat av upprepad stress och/eller kumulativt trauma på låg nivå, vilket leder till inflammation och sträckning av ledkapseln. Det vanligaste besväret är axiell ländryggssmärta med refererad smärta i flanken, höften och låret. Inga fysiska undersökningsfynd är patognomoniska för diagnos. Den starkaste indikatorn för lumbal facetogenisk smärta är smärtreduktion efter anestesiblock av rami mediales (mediala grenar) av rami dorsales som innerverar facettlederna. Eftersom falskt positiva och eventuellt falskt negativa resultat kan förekomma måste resultaten tolkas noggrant. Hos patienter med injektionsbekräftad zygapofysisk ledsmärta kan procedurinterventioner göras inom ramen för en multidisciplinär, multimodal behandlingsregim som inkluderar farmakoterapi, sjukgymnastik och regelbunden träning och, om så är indicerat, psykoterapi. För närvarande är guldstandarden för behandling av facetogen smärta radiofrekvensbehandling (1 B+). Bevisen som stöder intraartikulära kortikosteroider är begränsad; därför bör detta reserveras för dem som inte svarar på radiofrekvensbehandling (2 B1).
Facetogen smärta som härrör från ländryggsfasettlederna är en vanlig orsak till ländryggssmärta hos den vuxna befolkningen. Goldthwaite var den första som beskrev syndromet 1911, och Ghormley anses allmänt ha myntat termen �facetsyndrom� 1933. Facetogen smärta definieras som smärta som uppstår från vilken struktur som helst som är en del av facettlederna, inklusive den fibrösa kapseln , synovialmembran, hyalint brosk och ben.35
Vanligare är det resultatet av upprepad stress och/eller kumulativt trauma på låg nivå. Detta leder till inflammation, vilket kan göra att facettleden fylls med vätska och sväller, vilket resulterar i att ledkapseln töjs och efterföljande smärtgenerering.27 Inflammatoriska förändringar runt fasettledet kan också irritera ryggmärgen via försminalnedsättning, vilket leder till ischias. Dessutom fann Igarashi et al. 28 att inflammatoriska cytokiner som släpptes genom ventralledkapseln hos patienter med zygapofysial gemensamma degenerering kan vara delvis ansvariga för de neuropatiska symtomen hos individer med ryggmärgsstenos. Fördjupningsfaktorer för zygapofysial ledvärk innefattar spondylolistes / lysis, degenerativ skivjukdom och avancerad ålder. 5
IC ADDITIONAL PROV
Förekomsten av patologiska förändringar i facettlederna vid radiologisk undersökning beror på försökspersonernas medelålder, den använda radiologiska tekniken och definitionen av abnormitet. Degenerativa fasettfogar kan bäst visualiseras via beräknad tomografi (CT) undersökning. 49
NEUROPATISK SMÄRTA
Smärta initierad eller orsakad av en primär skada eller dysfunktion i det somatosensorära nervsystemet.
Neuropatisk smärta är vanligtvis kronisk, svår att behandla och ofta resistent mot vanliga smärtstillande behandlingar.
Abstrakt
Neuropatisk smärta orsakas av en skada eller sjukdom i det somatosensoriska systemet, inklusive perifera fibrer (Ap-, A- och C-fibrer) och centrala neuroner, och drabbar 7-10 % av den allmänna befolkningen. Flera orsaker till neuropatisk smärta har beskrivits. Dess förekomst kommer sannolikt att öka på grund av den åldrande globala befolkningen, ökad diabetes mellitus och förbättrad överlevnad från cancer efter kemoterapi. Obalanser mellan excitatorisk och hämmande somatosensorisk signalering, förändringar i jonkanaler och variation i hur smärtmeddelanden moduleras i det centrala nervsystemet har alla varit inblandade i neuropatisk smärta. Dessutom verkar bördan av kronisk neuropatisk smärta vara relaterad till komplexiteten av neuropatiska symtom, dåliga resultat och svåra behandlingsbeslut. Viktigt är att livskvaliteten försämras hos patienter med neuropatisk smärta på grund av ökade läkemedelsförskrivningar och besök hos vårdgivare och sjukligheten från själva smärtan och den anstiftande sjukdomen. Trots utmaningar sporrar framsteg i att förstå patofysiologin av neuropatisk smärta utvecklingen av nya diagnostiska procedurer och personliga ingrepp, som betonar behovet av ett multidisciplinärt tillvägagångssätt för hantering av neuropatisk smärta.
PATHOGENESIS AV NEUROPATISK PÄNN
PERIPHERAL MECHANISMS
Efter en perifer nervskada blir nervceller känsligare och utvecklar onormal excitabilitet och förhöjd känslighet för stimulering.
Detta är känt som ... Perifer sensibilisering!
CENTRALMEKANISMER
Som en konsekvens av pågående spontan aktivitet i periferin utvecklar neuroner en ökad bakgrundsaktivitet, förstorade receptiva fält och ökade svar på afferenta impulser, inklusive normala taktila stimuli. Detta är känt som ... Central Sensibilisering!
Kronisk neuropatisk smärta är vanligare hos kvinnor (8 % jämfört med 5.7 % hos män) och hos patienter >50 år (8.9 % mot 5.6 % hos de <49 år gamla), och drabbar oftast nedre delen av ryggen och nedre extremiteterna , hals och övre extremiteter24. Lumbala och cervikala smärtsamma radikulopatier är förmodligen den vanligaste orsaken till kronisk neuropatisk smärta. I överensstämmelse med dessa data visade en undersökning av >12,000 40 patienter med kronisk smärta med både nociceptiva och neuropatiska smärttyper, hänvisad till smärtspecialister i Tyskland, att 25 % av alla patienter upplevde åtminstone vissa egenskaper av neuropatisk smärta (såsom brännande känsla, domningar och stickningar); patienter med kronisk ryggsmärta och radikulopati drabbades särskiltXNUMX.
Klinisk neurofysiologins bidrag till förståelsen av huvudvärksmekanismerna av spännings-typ.
Abstrakt
Hittills har kliniska neurofysiologiska studier på spänningshuvudvärk (TTH) utförts med två huvudsyften: (1) för att fastställa om vissa neurofysiologiska parametrar kan fungera som markörer för TTH, och (2) för att undersöka fysiopatologin för TTH. När det gäller den första punkten är de nuvarande resultaten nedslående eftersom vissa avvikelser som hittas hos TTH-patienter också kan observeras ofta hos migränpatienter. Å andra sidan har klinisk neurofysiologi spelat en viktig roll i debatten om patogenesen av TTH. Studier av exteroceptiv undertryckning av temporalismuskelkontraktionen har upptäckt en dysfunktion i hjärnstammens excitabilitet och suprasegmentell kontroll. En liknande slutsats har nåtts med användning av trigeminocervikala reflexer, vars abnormiteter i TTH har antytt en minskad hämmande aktivitet av hjärnstammens interneuron, vilket återspeglar onormala endogena smärtkontrollmekanismer. Intressant nog verkar den neurala excitabilitetsabnormaliteten i TTH vara ett generaliserat fenomen, inte begränsat till kranialdistrikten. Defekta DNIC-liknande mekanismer har verkligen påvisats även i somatiska distrikt genom nociceptiva flexionsreflexstudier. Tyvärr är de flesta neurofysiologiska studier på TTH behäftade med allvarliga metodologiska brister, som bör undvikas i framtida forskning för att klargöra TTH-mekanismerna bättre.
Referenser:
Neurofysiologi av artritvärk. McDougall JJ1 Linton P.
Smärta härrörande från ländryggfasets leder. van Kleef M1, Vanelderen P, Cohen SP, Lataster A, Van Zundert J, Mekhail N.
Neuropatisk smärtaLuana Colloca,1Taylor Ludman,1Didier Bouhassira,2Ralf Baron,3Anthony H. Dickenson,4David Yarnitsky,5Roy Freeman,6Andrea Truini,7Nadine Attal, Nanna B. Finnerup,9Christopher Eccleston,1011Eija Kalso,12David L. Bennett,13Robert H. Dworkin,14och Srinivasa N. Raja15
Klinisk neurofysiologins bidrag till förståelsen av huvudvärksmekanismerna av spännings-typ. Rossi P1, Vollono C, Valeriani M, Sandrini G.
Läkare definierar kronisk smärta, som all smärta som varar i 3 till 6 månader eller mer. De smärta påverkar en individs psykiska hälsa och dagliga liv. Smärta kommer från en rad meddelanden som går genom nervsystemet. Depression verkar följa smärta. Det ger allvarliga symtom som påverkar hur en individ känner, tänker och hur han hanterar dagliga aktiviteter, det vill säga sova, äta och arbeta. Kiropraktor, Dr Alex Jimenez fördjupar sig i potentiella biomarkörer som kan hjälpa till att hitta och behandla grundorsakerna till smärta och kronisk smärta.
Det första steget i framgångsrik smärtbehandling är en omfattande biopsykosocial bedömning.
Omfattningen av organisk patologi kanske inte återspeglas exakt i smärtupplevelsen.
Den initiala bedömningen kan användas för att identifiera områden som kräver mer djupgående utvärdering.
Många validerade självrapporteringsverktyg finns tillgängliga för att bedöma effekten av kronisk smärta.
Bedömning av patienter med kronisk smärta
Kronisk smärta är ett folkhälsoproblem som drabbar 20–30 % av befolkningen i västerländska länder. Även om det har gjorts många vetenskapliga framsteg i förståelsen av smärtans neurofysiologi, är det inte enkelt eller väldefinierat att exakt bedöma och diagnostisera en patients kroniska smärtproblem. Hur kronisk smärta konceptualiseras påverkar hur smärta utvärderas och vilka faktorer som beaktas när man ställer en kronisk smärtdiagnos. Det finns inget en-till-en-samband mellan mängden eller typen av organisk patologi och smärtintensitet, utan istället, den kroniska smärtupplevelsen formas av en myriad av biomedicinska, psykosociala (t.ex. patienters övertygelse, förväntningar och humör) och beteendefaktorer (t.ex. sammanhang, reaktioner från betydande andra). Att bedöma var och en av dessa tre domäner genom en omfattande utvärdering av personen med kronisk smärta är avgörande för behandlingsbeslut och för att underlätta optimala resultat. Denna utvärdering bör innehålla en noggrann patienthistorik och medicinsk utvärdering och en kort screeningintervju där patientens beteende kan observeras. Ytterligare bedömning för att ta itu med frågor som identifierats under den första utvärderingen kommer att vägleda beslut om vilka ytterligare bedömningar, om några, som kan vara lämpliga. Standardiserade självrapporterade instrument för att utvärdera patientens smärtintensitet, funktionella förmågor, övertygelser och förväntningar samt känslomässigt lidande finns tillgängliga och kan administreras av läkaren, eller så kan en remiss för fördjupad utvärdering göras för att hjälpa till med behandlingsplanering.
Smärta är ett extremt utbrett symptom. Enbart kronisk smärta beräknas påverka 30 % av den vuxna befolkningen i USA, uppemot 100 miljoner vuxna.1
Trots den skyhöga kostnaden för att behandla människor med kronisk smärta, är lindring för många fortfarande svårfångad och fullständig eliminering av smärta är sällsynt. Även om det har gjorts betydande framsteg i kunskapen om smärtans neurofysiologi, tillsammans med utvecklingen av potenta smärtstillande mediciner och andra innovativa medicinska och kirurgiska ingrepp, är i genomsnitt mängden smärtreduktion med tillgängliga procedurer 30–40 % och detta inträffar i färre än hälften av de behandlade patienterna.
Sättet vi tänker på smärta påverkar hur vi utvärderar smärta. Bedömningen börjar med anamnes och fysisk undersökning, följt av laboratorietester och bilddiagnostiska procedurer i ett försök att identifiera och/eller bekräfta förekomsten av någon underliggande patologi som orsakar symtomen/symptomen eller smärtgenerator.
I avsaknad av identifierbar organisk patologi kan vårdgivaren anta att rapporten om symtom härrör från psykologiska faktorer och kan begära en psykologisk utvärdering för att upptäcka de känslomässiga faktorer som ligger till grund för patientens rapport. Det finns dualitet där rapporten om symtom tillskrivs antingen somatisk or psykogena mekanismer.
Som ett exempel, de organiska baserna för några av de vanligaste och återkommande akuta (t.ex. huvudvärk)3 och kroniska [t.ex. ryggsmärta, fibromyalgi (FM)] smärtproblem är i stort sett okända,4,5 medan å andra sidan asymtomatiska individer kan ha strukturella avvikelser såsom diskbråck som skulle förklara smärta om den fanns.6,7�Det saknas adekvata förklaringar för patienter utan identifierad organisk patologi som rapporterar svår smärta och smärtfria individer med betydande, objektiv patologi.
Kronisk smärta påverkar mer än bara den enskilde patienten, utan även dennes betydelsefulla andra (partners, släktingar, arbetsgivare och arbetskamrater och vänner), vilket gör lämplig behandling nödvändig. Tillfredsställande behandling kan endast komma från en omfattande bedömning av smärtans biologiska etiologi i samband med patientens specifika psykosociala och beteendemässiga presentation, inklusive deras känslomässiga tillstånd (t.ex. ångest, depression och ilska), uppfattning och förståelse av symtom och reaktioner på dessa. symptom från betydande andra.8,9 En nyckelpremiss är att flera faktorer påverkar symptomen och funktionella begränsningar hos individer med kronisk smärta. Därför behövs en omfattande bedömning som tar upp biomedicinska, psykosociala och beteendemässiga domäner, eftersom var och en bidrar till kronisk smärta och relaterad funktionsnedsättning.10,11
Omfattande bedömning av en individ med kronisk smärta
Turk och Meichenbaum12 föreslog att tre centrala frågor bör vägleda bedömningen av personer som rapporterar smärta:
Hur stor är omfattningen av patientens sjukdom eller skada (fysisk funktionsnedsättning)?
Hur stor är sjukdomen? Det vill säga, i vilken utsträckning lider patienten, är handikappad och kan inte njuta av vanliga aktiviteter?
Verkar individens beteende lämpligt för sjukdomen eller skadan, eller finns det några tecken på symtomförstärkning av någon av en mängd olika psykologiska eller sociala skäl (t.ex. fördelar som positiv uppmärksamhet, humörförändrande mediciner, ekonomisk kompensation)?
För att besvara dessa frågor bör information samlas in från patienten genom anamnes och fysisk undersökning, i kombination med en klinisk intervju och genom standardiserade bedömningsinstrument. Sjukvårdsleverantörer måste söka alla orsaker till smärta genom fysisk undersökning och diagnostiska tester samtidigt som de utvärderar patientens humör, rädslor, förväntningar, hanteringsinsatser, resurser, reaktioner från betydande andra och smärtans inverkan på patienterna� liv.11 Kort sagt måste vårdgivaren utvärdera �hela personen� och inte bara smärtan.
De allmänna målen för historien och medicinsk utvärdering är att:
(i) fastställa behovet av ytterligare diagnostiska tester
(ii) avgöra om medicinska data kan förklara patientens symtom, symtoms svårighetsgrad och funktionella begränsningar
(iii) ställa en medicinsk diagnos
(iv) utvärdera tillgången på lämplig behandling
v) fastställa målen för behandlingen
(vi) bestämma lämplig kurs för symtomhantering om ett fullständigt botemedel inte är möjligt.
Ett betydande antal patienter som rapporterar kronisk smärta visar ingen fysisk patologi med vanliga röntgenbilder, datoriserade axialtomografi eller elektromyografi (en omfattande litteratur finns tillgänglig om fysisk bedömning, röntgen- och laboratoriebedömningsprocedurer för att fastställa den fysiska grunden för smärta),17 vilket gör en exakt patologisk diagnos svår eller omöjlig.
Trots dessa begränsningar förblir patientens anamnes och fysiska undersökning basen för medicinsk diagnos, kan ge ett skydd mot övertolkning av fynd från diagnostisk bilddiagnostik som till stor del är bekräftande, och kan användas för att styra riktningen för ytterligare utvärderingsinsatser.
Dessutom konsumerar patienter med kroniska smärtproblem ofta en mängd olika mediciner.18 Det är viktigt att diskutera en patients aktuella mediciner under intervjun, eftersom många smärtstillande mediciner är förknippade med biverkningar som kan orsaka eller efterlikna känslomässigt lidande.19 Vårdgivare bör inte bara känna till mediciner som används för kronisk smärta, utan också med biverkningar från dessa mediciner som resulterar i trötthet, sömnsvårigheter och humörförändringar för att undvika feldiagnostik av depression.
Användningen av dagliga dagböcker tros vara mer exakt eftersom de är baserade på realtid snarare än återkallande. Patienter kan bli ombedda att föra regelbundna dagböcker över smärtintensitet med värderingar registrerade flera gånger varje dag (t.ex. måltider och läggdags) under flera dagar eller veckor och flera smärtvärden kan beräknas i genomsnitt över tiden.
Ett problem som noterats med användningen av dagböcker med papper och penna är att patienter kanske inte följer instruktionerna för att ge betyg vid specificerade intervall. Snarare kan patienter fylla i dagböcker i förväg (�fyll framåt�) eller strax innan de träffar en läkare (�fyll baklänges�),24 vilket undergräver dagböckernas förmodade giltighet. Elektroniska dagböcker har vunnit acceptans i vissa forskningsstudier för att undvika dessa problem.
Forskning har visat vikten av att bedöma övergripande hälsorelaterad livskvalitet (HRQOL) hos patienter med kronisk smärta förutom funktion.31,32 Det finns ett antal väletablerade, psykometriskt stödda HRQOL-mått [Medical Outcomes Study Short-Form Health Survey (SF-36)],33 allmänna mått på fysisk funktion [t.ex. Pain Disability Index (PDI)],34 och sjukdomsspecifika mått [t.ex. Western Ontario MacMaster Osteoarthritis Index (WOMAC);35 Roland-Morris Back Pain Disability Questionnaire (RDQ) )]36 för att bedöma funktion och livskvalitet.
Sjukdomsspecifika mått är utformade för att utvärdera effekten av ett specifikt tillstånd (t.ex. smärta och stelhet hos personer med artros), medan generiska mått gör det möjligt att jämföra fysisk funktion förknippad med en viss sjukdom och dess behandling med olika andra tillstånd. Specifika effekter av en störning kanske inte upptäcks när ett generiskt mått används; därför kan sjukdomsspecifika åtgärder vara mer benägna att avslöja kliniskt viktig förbättring eller försämring av specifika funktioner som ett resultat av behandling. Allmänna mått på funktion kan vara användbara för att jämföra patienter med en mångfald av smärtsamma tillstånd. Den kombinerade användningen av sjukdomsspecifika och generiska åtgärder underlättar uppnåendet av båda målen.
Förekomsten av känslomässig ångest hos personer med kronisk smärta utgör en utmaning när man ska bedöma symtom som trötthet, minskad aktivitetsnivå, minskad libido, aptitförändring, sömnstörningar, viktökning eller viktminskning samt minnes- och koncentrationsbrist, eftersom dessa symtom kan vara resultat av smärta, känslomässigt lidande eller behandlingsmediciner som ordinerats för att kontrollera smärta.
Instrument har utvecklats specifikt för smärtpatienter för att bedöma psykisk ångest, smärtans inverkan på patienternas liv, känsla av kontroll, hanteringsbeteenden och attityder om sjukdom, smärta och vårdgivare.17
Till exempel är Beck Depression Inventory (BDI)39 och Profile of Mood States (POMS)40 psykometriskt bra för att bedöma symtom på deprimerat humör, känslomässigt lidande och humörstörningar, och har rekommenderats att användas i alla kliniska prövningar av kronisk smärta;41 poängen måste dock tolkas med försiktighet och kriterierna för nivåer av känslomässigt lidande kan behöva modifieras för att förhindra falska positiva resultat.42
Labbiomarkörer för smärta
Biomarkörer är biologiska egenskaper som kan användas för att indikera hälsa eller sjukdom. Denna artikel granskar studier av biomarkörer för ländryggssmärta (LBP) hos människor. LBP är den främsta orsaken till funktionshinder, orsakad av olika ryggradsrelaterade störningar, inklusive intervertebral diskdegeneration, diskbråck, spinal stenos och facettartrit. Fokus för dessa studier är inflammatoriska mediatorer, eftersom inflammation bidrar till patogenesen av diskdegeneration och associerade smärtmekanismer. Studier tyder i allt högre grad på att förekomsten av inflammatoriska mediatorer kan mätas systemiskt i blodet. Dessa biomarkörer kan fungera som nya verktyg för att styra patientvården. För närvarande är patientens svar på behandlingen oförutsägbar med en betydande återfallsfrekvens, och även om kirurgiska behandlingar kan ge anatomisk korrigering och smärtlindring, är de invasiva och kostsamma. Översikten omfattar studier utförda på populationer med specifika diagnoser och odefinierat ursprung till LBP. Eftersom den naturliga historien för LBP är progressiv, kategoriseras studiernas tidsmässiga karaktär efter varaktigheten av symtomologi/sjukdom. Relaterade studier om förändringar i biomarkörer med behandling granskas också. I slutändan har diagnostiska biomarkörer för LBP och ryggradsdegeneration potentialen att skapa en era av individualiserad ryggradsmedicin för personlig terapi vid behandling av LBP.
Biomarkörer för kronisk neuropatisk smärta och potentiell tillämpning vid ryggmärgsstimulering
Denna recension fokuserade på att förstå vilka ämnen i människokroppen som ökar och minskar med ökande neuropatisk smärta. Vi granskade olika studier och såg samband mellan neuropatisk smärta och komponenter i immunsystemet (det här systemet försvarar kroppen mot sjukdomar och infektioner). Våra fynd kommer särskilt att vara användbara för att förstå sätt att minska eller eliminera det obehag som kronisk neuropatisk smärta för med sig. Ryggmärgsstimulering (SCS) procedur är en av få ganska effektiva behandlingar för smärtlindring. En uppföljningsstudie kommer att tillämpa våra resultat från denna översyn på SCS, för att förstå mekanismen och ytterligare optimera effektiviteten.
Pro-inflammatoriska cytokiner såsom IL-1?, IL-6, IL-2, IL-33, CCL3, CXCL1, CCR5 och TNF-a, har visat sig spela betydande roller i amplifieringen av kroniska smärttillstånd.
Efter genomgång av olika studier relaterade till smärtbiomarkörer fann vi att serumnivåer av pro-inflammatoriska cytokiner och kemokiner, såsom IL-1?, IL-6, IL-2, IL-33, CCL3, CXCL1, CCR5 och TNF -?, var signifikant uppreglerade under kronisk smärtupplevelse. Å andra sidan visade sig antiinflammatoriska cytokiner som IL-10 och IL-4 visa signifikant nedreglering under kroniskt smärttillstånd.
Biomarkörer för depression
En uppsjö av forskning har involverat hundratals förmodade biomarkörer för depression, men har ännu inte helt klarlagt deras roller i depressiv sjukdom eller fastställt vad som är onormalt hos vilka patienter och hur biologisk information kan användas för att förbättra diagnos, behandling och prognos. Denna brist på framsteg beror delvis på depressionens natur och heterogenitet, i kombination med metodologisk heterogenitet inom forskningslitteraturen och det stora utbudet av biomarkörer med potential, vars uttryck ofta varierar beroende på många faktorer. Vi granskar den tillgängliga litteraturen, som indikerar att markörer involverade i inflammatoriska, neurotrofiska och metaboliska processer, såväl som neurotransmittor och neuroendokrina systemkomponenter, representerar mycket lovande kandidater. Dessa kan mätas genom genetiska och epigenetiska, transkriptomiska och proteomiska, metabolomiska och neuroavbildningsbedömningar. Användningen av nya tillvägagångssätt och systematiska forskningsprogram krävs nu för att avgöra om och vilka biomarkörer som kan användas för att förutsäga svar på behandling, stratifiera patienter till specifika behandlingar och utveckla mål för nya insatser. Vi drar slutsatsen att det finns mycket lovande för att minska bördan av depression genom att vidareutveckla och utöka dessa forskningsvägar.
Referenser:
Bedömning av patienter med kronisk smärta�EJ Dansiet och DC Turk*t�
Inflammatoriska biomarkörer för ländryggssmärta och diskdegeneration: en recension.
Khan AN1, Jacobsen HE2, Khan J1, Filippi CG3, Levine M3, Lehman RA Jr2,4, Riew KD2,4, Lenke LG2,4, Chahine NO2,5.
Biomarkörer för kronisk neuropatisk smärta och deras potentiella tillämpning vid ryggmärgsstimulering: en översyn
Chibueze D. Nwagwu,1 Christina Sarris, MD,3 Yuan-Xiang Tao, Ph.D., MD,2 och Antonios Mammis, MD1,2
Biomarkörer för depression: nya insikter, nuvarande utmaningar och framtidsutsikter. Strawbridge R1, Young AH1,2, Cleare AJ1,2.
Smärta är den mänskliga kroppens naturliga reaktion på skada eller sjukdom, och det är ofta en varning om att något är fel. När problemet är löst slutar vi i allmänhet uppleva dessa smärtsamma symtom, men vad händer när smärtan fortsätter långt efter att orsaken är borta? Kronisk smärta är medicinskt definierad som ihållande smärta som varar i 3 till 6 månader eller mer. Kronisk smärta är förvisso ett utmanande tillstånd att leva med, som påverkar allt från individens aktivitetsnivå och arbetsförmåga samt deras personliga relationer och psykologiska tillstånd. Men är du medveten om att kronisk smärta också kan påverka din hjärnas struktur och funktion? Det visar sig att dessa hjärnförändringar kan leda till både kognitiv och psykologisk funktionsnedsättning.
Kronisk smärta påverkar inte bara en enskild del av sinnet, den kan faktiskt resultera i förändringar i många viktiga områden i hjärnan, varav de flesta är involverade i många grundläggande processer och funktioner. Olika forskningsstudier genom åren har funnit förändringar i hippocampus, tillsammans med minskning av grå substans från den dorsolaterala prefrontala cortex, amygdala, hjärnstammen och höger insulär cortex, för att nämna några, förknippade med kronisk smärta. En uppdelning av några av strukturerna i dessa regioner och deras relaterade funktioner kan hjälpa till att sätta dessa hjärnförändringar i ett sammanhang, för många individer med kronisk smärta. Syftet med följande artikel är att demonstrera och diskutera de strukturella och funktionella förändringar i hjärnan som är förknippade med kronisk smärta, särskilt i de fall där dessa förmodligen varken återspeglar skada eller atrofi.
Strukturella hjärnförändringar i kronisk smärta återspeglar förmodligen varken skada eller atrofi
Abstrakt
Kronisk smärta verkar vara associerad med minskning av hjärnans grå substans i områden som kan tillskrivas överföringen av smärta. De morfologiska processerna som ligger bakom dessa strukturella förändringar, troligen efter funktionell omorganisation och central plasticitet i hjärnan, är fortfarande oklara. Smärtan vid höftledsartros är ett av få kroniska smärtsyndrom som i huvudsak går att bota. Vi undersökte 20 patienter med kronisk smärta på grund av unilateral coxarthrosis (medelålder 63.25–9.46 (SD) år, 10 kvinnor) före endoprotetisk höftledsoperation (smärttillstånd) och övervakade hjärnstrukturella förändringar upp till 1 år efter operationen: 6–8 veckor , 12–18 veckor och 10–14 månader när det är helt smärtfritt. Patienter med kronisk smärta på grund av unilateral coxarthrosis hade signifikant mindre grå substans jämfört med kontroller i främre cingulate cortex (ACC), insular cortex och operculum, dorsolateral prefrontal cortex (DLPFC) och orbitofrontal cortex. Dessa regioner fungerar som multiintegrativa strukturer under upplevelsen och förväntan av smärta. När patienterna var smärtfria efter återhämtning från endoproteskirurgi, upptäcktes en ökning av grå substans i nästan samma områden. Vi fann också en progressiv ökning av hjärnans grå substans i den premotoriska cortexen och det kompletterande motoriska området (SMA). Vi drar slutsatsen att avvikelser i grå substans vid kronisk smärta inte är orsaken, utan sekundära till sjukdomen och åtminstone delvis beror på förändringar i motorisk funktion och kroppslig integration.
Beskrivning
Bevis på funktionell och strukturell omorganisation hos patienter med kronisk smärta stöder idén att kronisk smärta inte bara bör begreppsliggöras som ett förändrat funktionstillstånd, utan också som en konsekvens av funktionell och strukturell hjärnplasticitet [1], [2], [3], [4], [5], [6]. Under de senaste sex åren har mer än 20 studier publicerats som visar strukturella förändringar i hjärnan i 14 kroniska smärtsyndrom. Ett slående särdrag i alla dessa studier är det faktum att förändringarna i den grå substansen inte var slumpmässigt fördelade, utan förekommer i definierade och funktionellt mycket specifika hjärnområden – nämligen involvering i supraspinal nociceptiv bearbetning. De mest framträdande fynden var olika för varje smärtsyndrom, men överlappade i cingulate cortex, orbitofrontal cortex, insula och dorsala pons [4]. Ytterligare strukturer omfattar thalamus, dorsolateral prefrontal cortex, basala ganglier och hippocampusområdet. Dessa fynd diskuteras ofta som cellulär atrofi, vilket förstärker idén om skada eller förlust av hjärnans grå substans [7], [8], [9]. Faktum är att forskare fann ett samband mellan minskningar av hjärnans grå substans och varaktigheten av smärta [6], [10]. Men smärtans varaktighet är också kopplad till patientens ålder, och den åldersberoende globala, men även regionalt specifika nedgången av grå substans är väl dokumenterad [11]. Å andra sidan kan dessa strukturella förändringar också vara en minskning av cellstorlek, extracellulära vätskor, synaptogenes, angiogenes eller till och med på grund av blodvolymförändringar [4], [12], [13]. Oavsett källan är det för vår tolkning av sådana fynd viktigt att se dessa morfometriska fynd i ljuset av en mängd morfometriska studier i träningsberoende plasticitet, givet att regionalt specifika strukturella hjärnförändringar har visats upprepade gånger efter kognitiv och fysisk träning [ 14].
Det är inte förstått varför endast en relativt liten del av människor utvecklar ett kroniskt smärtsyndrom, med tanke på att smärta är en universell upplevelse. Frågan uppstår om hos vissa människor en strukturell skillnad i centrala smärtöverförande system kan fungera som en diatese för kronisk smärta. Grå substansförändringar i fantomsmärta på grund av amputation [15] och ryggmärgsskada [3] tyder på att de morfologiska förändringarna i hjärnan, åtminstone delvis, är en konsekvens av kronisk smärta. Smärtan vid höftledsartros (OA) är dock ett av få kroniska smärtsyndrom som huvudsakligen kan botas, eftersom 88 % av dessa patienter regelbundet är fria från smärta efter total höftprotesoperation (THR) [16]. I en pilotstudie har vi analyserat tio patienter med höft-OA före och kort efter operationen. Vi fann minskningar av grå substans i den främre cingulerade cortex (ACC) och insula under kronisk smärta före THR-operation och fann ökningar av grå substans i motsvarande hjärnområden i smärtfritt tillstånd efter operationen [17]. Med fokus på detta resultat utökade vi nu våra studier som undersökte fler patienter (n?=?20) efter framgångsrik THR och övervakade strukturella förändringar i hjärnan i fyra tidsintervall, upp till ett år efter operationen. För att kontrollera för förändringar i grå substans på grund av motorisk förbättring eller depression administrerade vi också frågeformulär inriktade på förbättring av motorisk funktion och mental hälsa.
Material och metoder
Frivilliga
Patienterna som rapporteras här är en undergrupp av 20 patienter av 32 nyligen publicerade patienter som jämfördes med en ålders- och könsmatchad frisk kontrollgrupp [17] men deltog i ytterligare ett års uppföljningsundersökning. Efter operationen hoppade 12 patienter av på grund av en andra endoprotetisk operation (n?=?2), allvarlig sjukdom (n?=?2) och återkallande av samtycke (n?=?8). Detta lämnade en grupp på tjugo patienter med unilateral primär höft-OA (medelålder 63.25–9.46 (SD) år, 10 kvinnor) som undersöktes fyra gånger: före operation (smärttillstånd) och igen 6–8 och 12–18 veckor och 10 �14 månader efter endoproteskirurgi, när den är helt smärtfri. Alla patienter med primär höft-OA hade en smärtanamnes längre än 12 månader, från 1 till 33 år (medelvärde 7.35 år) och ett genomsnittligt smärtpoäng på 65.5 (från 40 till 90) på en visuell analog skala (VAS) från 0 (ingen smärta) till 100 (värsta tänkbara smärta). Vi utvärderade varje förekomst av mindre smärthändelser, inklusive tand-, öron- och huvudvärk upp till 4 veckor före studien. Vi valde också slumpmässigt data från 20 köns- och åldersmatchade friska kontroller (medelålder 60,95–8,52 (SD) år, 10 kvinnor) av de 32 i ovan nämnda pilotstudie [17]. Ingen av de 20 patienterna eller av de 20 köns- och åldersmatchade friska frivilliga hade någon neurologisk eller intern medicinsk historia. Studien fick etiskt godkännande av den lokala etiska kommittén och skriftligt informerat samtycke erhölls från alla studiedeltagare innan undersökningen.
Beteendedata
Vi samlade in data om depression, somatisering, ångest, smärta och fysisk och mental hälsa hos alla patienter och alla fyra tidpunkter med hjälp av följande standardiserade frågeformulär: Beck Depression Inventory (BDI) [18], Brief Symptom Inventory (BSI) [19], Schmerzempfindungs-Skala (SES?=?smärta obehagsskala) [20] och Health Survey 36-Item Short Form (SF-36) [21] och Nottingham Health Profile (NHP). Vi genomförde upprepade åtgärder ANOVA och parade tvåsidiga t-tester för att analysera longitudinella beteendedata med SPSS 13.0 för Windows (SPSS Inc., Chicago, IL) och använde Greenhouse Geisser-korrigering om antagandet om sfäricitet kränktes. Signifikansnivån sattes till p<0.05.
VBM – Datainsamling
Bildförvärv. Högupplöst MR-skanning utfördes på ett 3T MRI-system (Siemens Trio) med en standard 12-kanals huvudspole. För var och en av de fyra tidpunkterna, skanning I (mellan 1 dag och 3 månader före endoproteskirurgi), skanning II (6 till 8 veckor efter operationen), skanning III (12 till 18 veckor efter operationen) och skanning IV (10–14) månader efter operationen) förvärvades en T1-vägd strukturell MRT för varje patient med en 3D-FLASH-sekvens (TR 15 ms, TE 4.9 ms, vändvinkel 25�, 1 mm skivor, FOV 256�256, voxelstorlek 1�1� 1 mm).
Bildbehandling och statistisk analys
Dataförbehandling och analys utfördes med SPM2 (Wellcome Department of Cognitive Neurology, London, Storbritannien) som kördes under Matlab (Mathworks, Sherborn, MA, USA) och innehöll en voxelbaserad morfometri (VBM)-verktygslåda för longitudinella data, som baseras på högupplösta strukturella 3D MR-bilder och gör det möjligt att tillämpa voxel-vis statistik för att upptäcka regionala skillnader i grå substans densitet eller volymer [22], [23]. Sammanfattningsvis involverade förbearbetning rumslig normalisering, segmentering av grå substans och 10 mm spatial utjämning med en Gaussisk kärna. För förbehandlingsstegen använde vi ett optimerat protokoll [22], [23] och en skanner- och studiespecifik grå substansmall [17]. Vi använde SPM2 snarare än SPM5 eller SPM8 för att göra denna analys jämförbar med vår pilotstudie [17]. eftersom det tillåter en utmärkt normalisering och segmentering av longitudinella data. Men eftersom en nyare uppdatering av VBM (VBM8) nyligen blev tillgänglig (dbm.neuro.uni-jena.de/vbm/), använde vi också VBM8.
Tvärsnittsanalys
Vi använde ett t-test med två prov för att upptäcka regionala skillnader i hjärnans grå substans mellan grupper (patienter vid tidpunktsskanning I (kronisk smärta) och friska kontroller). Vi tillämpade ett tröskelvärde på p<0.001 (okorrigerat) över hela hjärnan på grund av vår starka a priory-hypotes, som är baserad på 9 oberoende studier och kohorter som visar minskningar av grå substans hos patienter med kronisk smärta [7], [8], [ 9], [15], [24], [25], [26], [27], [28], att ökningar av grå substans kommer att dyka upp i samma (för smärtbehandlingsrelevanta) regioner som i vår pilotstudie (17) ). Grupperna matchades för ålder och kön utan några signifikanta skillnader mellan grupperna. För att undersöka om skillnaderna mellan grupperna förändrades efter ett år jämförde vi även patienter vid tidpunktsskanning IV (smärtfri, ett års uppföljning) med vår friska kontrollgrupp.
Longitudinell analys
För att upptäcka skillnader mellan tidpunkter (Scan I�IV) jämförde vi skanningarna före operationen (smärttillstånd) och igen 6�8 och 12�18 veckor och 10�14 månader efter endoproteskirurgi (smärtfri) som upprepad åtgärd ANOVA. Eftersom eventuella förändringar i hjärnan på grund av kronisk smärta kan behöva lite tid för att avta efter operation och upphörande av smärta och på grund av smärtan efter operationen som patienterna rapporterade, jämförde vi i longitudinell analys scan I och II med scan III och IV. För att upptäcka förändringar som inte är nära kopplade till smärta letade vi också efter progressiva förändringar över alla tidsintervall. Vi vände hjärnan på patienter med OA i vänster höft (n?=?7) för att normalisera för sidan av smärtan för båda, gruppjämförelsen och longitudinell analys, men analyserade i första hand de oförändrade data. Vi använde BDI-poängen som en kovariat i modellen.
Resultat
Beteendeuppgifter
Alla patienter rapporterade kronisk höftsmärta före operationen och var smärtfria (avseende denna kroniska smärta) direkt efter operationen, men rapporterade ganska akut smärta efter operationen på scan II som skilde sig från smärtan på grund av artros. Den mentala hälsopoängen för SF-36 (F(1.925/17.322)?=?0.352, p?=?0.7) och BSI globala poäng GSI (F(1.706/27.302)?=?3.189, p?=?0.064 ) visade inga förändringar under tidsförloppet och ingen mental komorbiditet. Ingen av kontrollerna rapporterade någon akut eller kronisk smärta och ingen visade några symtom på depression eller fysisk/psykisk funktionsnedsättning.
Före operationen visade vissa patienter milda till måttliga depressiva symtom i BDI-poäng som minskade signifikant på skanning III (t(17)?=?2.317, p?=?0.033) och IV (t(16)?=?2.132, p? =?0.049). Dessutom förbättrades SES-poängen (smärta obehag) för alla patienter signifikant från scan I (före operationen) till scan II (t(16)?=?4.676, p<0.001), skanning III (t(14)?=? 4.760, p<0.001) och scan IV (t(14)?=?4.981, p<0.001, 1 år efter operationen) då smärtobehag minskade med smärtintensiteten. Smärtvärderingen på skanning 1 och 2 var positiv, samma betyg på dag 3 och 4 negativ. SES beskriver bara kvaliteten på upplevd smärta. Det var därför positivt på dag 1 och 2 (medelvärde 19.6 på dag 1 och 13.5 på dag 2) och negativt (na) på dag 3 och 4. Vissa patienter förstod dock inte denna procedur och använde SES som en global kvalitet av livet� mått. Det är därför alla patienter tillfrågades samma dag individuellt och av samma person om smärtförekomst.
I den korta hälsoundersökningen (SF-36), som består av de sammanfattande måtten av ett fysiskt hälsopoäng och ett mentalt hälsopoäng [29], förbättrades patienterna signifikant i fysisk hälsopoäng från skanning I till skanning II (t( 17)?=??4.266, p?=?0.001), scan III (t(16)?=??8.584, p<0.001) och IV (t(12)?=??7.148, p<0.001), men inte i den mentala hälsopoängen. Resultaten av NHP var liknande, i subskalan �smärta� (omvänd polaritet) observerade vi en signifikant förändring från scan I till scan II (t(14)?=??5.674, p<0.001, scan III (t(12) )?=??7.040, p<0.001 och skanning IV (t(10)?=??3.258, p?=?0.009). Vi fann också en signifikant ökning av subskalan �fysisk rörlighet� från scan I till scan III (t(12)?=??3.974, p?=?0.002) och scan IV (t(10)?=??2.511, p?=?0.031). Det fanns ingen signifikant förändring mellan scan I och scan II ( sex veckor efter operationen).
Strukturella data
Tvärsnittsanalys. Vi inkluderade ålder som en kovariat i den allmänna linjära modellen och hittade inga åldersförändringar. Jämfört med köns- och åldersmatchade kontroller, visade patienter med primär höft-OA (n?=?20) preoperativt (Scan I) minskad grå substans i främre cingulate cortex (ACC), insular cortex, operculum, dorsolateral prefrontal cortex ( DLPFC), höger temporal pol och lillhjärnan (tabell 1 och figur 1). Förutom höger putamen (x?=?31, y?=??14, z?=??1; p<0.001, t?=?3.32) hittades ingen signifikant ökning av densitet av grå substans hos patienter med OA jämfört med till friska kontroller. Vid jämförelse av patienter vid tidpunktsskanning IV med matchade kontroller hittades samma resultat som i tvärsnittsanalysen med användning av skanning I jämfört med kontroller.
Figur 1: Statistiska parametriska kartor som visar de strukturella skillnaderna i grå substans hos patienter med kronisk smärta på grund av primär höft-OA jämfört med kontroller och longitudinellt jämfört med dem själva över tid. Signifikanta förändringar i grå substans visas överlagrade i färg, tvärsnittsdata är avbildade i rött och longitudinella data i gult. Axialplan: den vänstra sidan av bilden är den vänstra sidan av hjärnan. top: Områden med signifikant minskning av grå substans mellan patienter med kronisk smärta på grund av primär höft-OA och opåverkade kontrollpersoner. p<0.001 okorrigerad botten: Ökning av grå substans hos 20 smärtfria patienter vid den tredje och fjärde skanningsperioden efter total höftprotesoperation, jämfört med den första (preoperativa) och andra (6–8 veckor efter operationen) skanningen. p<0.001 okorrigerade plots: Kontrastuppskattningar och 90 % konfidensintervall, effekter av intresse, godtyckliga enheter. x-axel: kontraster för de 4 tidpunkterna, y-axel: kontrastuppskattning vid ?3, 50, 2 för ACC och kontrastuppskattning vid 36, 39, 3 för insula.
Att vända data från patienter med vänster höft-OA (n?=?7) och jämföra dem med friska kontroller förändrade inte resultaten signifikant, men för en minskning av thalamus (x?=?10, y?=??20, z?=?3, p<0.001, t?=?3.44) och en ökning av höger lillhjärna (x?=?25, y?=??37, z?=??50, p<0.001, t? =?5.12) som inte nådde signifikans i patienternas oförändrade data jämfört med kontroller.
Longitudinell analys. I den longitudinella analysen upptäcktes en signifikant ökning (p<.001 okorrigerad) av grå substans genom att jämföra den första och andra skanningen (kronisk smärta/smärta efter operation) med den tredje och fjärde skanningen (smärtfri) i ACC, insular cortex, cerebellum och pars orbitalis hos patienter med OA (tabell 2 och figur 1). Grå substans minskade med tiden (p<001 helhjärnanalys okorrigerad) i den sekundära somatosensoriska cortexen, hippocampus, midcingulate cortex, thalamus och caudate nucleus hos patienter med OA (Figur 2).
Figur 2: a) Betydande ökningar av hjärnans grå substans efter framgångsrik operation. Axial vy av signifikant minskning av grå substans hos patienter med kronisk smärta på grund av primär höft-OA jämfört med kontrollpersoner. p<0.001 okorrigerad (tvärsnittsanalys), b) Longitudinell ökning av grå substans över tid i gult jämförande scan I&IIscan III>scan IV) hos patienter med OA. p<0.001 okorrigerad (longitudinell analys). Den vänstra sidan av bilden är den vänstra sidan av hjärnan.
Att vända data från patienter med vänster höft-OA (n?=?7) förändrade inte resultaten signifikant, men för en minskning av hjärnans grå substans i Heschls Gyrus (x?=??41, y?=?? 21, z?=?10, p<0.001, t?=?3.69) och Precuneus (x?=?15, y?=??36, z?=?3, p<0.001, t?=?4.60) .
Genom att kontrastera den första skanningen (förkirurgi) med skanning 3+4 (postkirurgi) fann vi en ökning av grå substans i frontal cortex och motorisk cortex (p<0.001 okorrigerad). Vi noterar att denna kontrast är mindre sträng eftersom vi nu har färre skanningar per tillstånd (smärta vs. icke-smärta). När vi sänker tröskeln upprepar vi det vi har hittat med kontrasten 1+2 mot 3+4.
Genom att leta efter områden som ökar över alla tidsintervall, fann vi förändringar av hjärnans grå substans i motoriska områden (område 6) hos patienter med coxarthrosis efter total höftprotes (scan Idbm.neuro.uni-jena.de/vbm/) vi kunde replikera detta fynd i den främre och mitten av cingulate cortex och båda främre insulae.
Vi beräknade effektstorlekarna och tvärsnittsanalysen (patienter vs. kontroller) gav en Cohen�sd på 1.78751 i toppvoxeln för ACC (x?=??12, y?=?25, z?=?? 16). Vi beräknade också Cohen�sd för den longitudinella analysen (kontrastskanning 1+2 vs. skanning 3+4). Detta resulterade i en Cohen�sd på 1.1158 i ACC (x?=??3, y?=?50, z?=?2). När det gäller ön (x?=??33, y?=?21, z?=?13) och relaterad till samma kontrast, är Cohen�sd 1.0949. Dessutom beräknade vi medelvärdet av icke-noll voxelvärden för Cohensd-kartan inom ROI (bestående av den främre uppdelningen av cingulate gyrus och subcallosal cortex, härledd från Harvard-Oxford Cortical Structural Atlas): 1.251223.
Dr Alex Jimenez insikt
Patienter med kronisk smärta kan uppleva en mängd olika hälsoproblem över tiden, bortsett från deras redan försvagande symtom. Till exempel kommer många individer att uppleva sömnproblem som ett resultat av sin smärta, men viktigast av allt, kronisk smärta kan också leda till olika psykiska problem, inklusive ångest och depression. Effekterna som smärta kan ha på hjärnan kan verka alltför överväldigande men växande bevis tyder på att dessa förändringar i hjärnan inte är permanenta och kan vändas när patienter med kronisk smärta får rätt behandling för sina underliggande hälsoproblem. Enligt artikeln återspeglar avvikelser i grå substans som hittas vid kronisk smärta inte hjärnskador, utan snarare är de en reversibel konsekvens som normaliseras när smärtan behandlas adekvat. Lyckligtvis finns en mängd olika behandlingsmetoder tillgängliga för att lindra kroniska smärtsymptom och återställa hjärnans struktur och funktion.
Diskussion
Genom att övervaka hela hjärnans struktur över tid bekräftar och utökar vi våra pilotdata som nyligen publicerats [17]. Vi fann förändringar i hjärnans grå substans hos patienter med primär höftledsartros i kroniskt smärttillstånd, som delvis vänder när dessa patienter är smärtfria efter endoproteskirurgi i höftleden. Den partiella ökningen av grå substans efter operationen är nästan i samma områden där en minskning av grå substans har setts före operationen. Att vända data från patienter med vänster höft-OA (och därför normalisera för sidan av smärtan) hade endast liten inverkan på resultaten men visade dessutom en minskning av grå substans i Heschls gyrus och Precuneus som vi inte enkelt kan förklara och, eftersom ingen a priori hypotes existerar, betrakta med stor försiktighet. Skillnaden som sågs mellan patienter och friska kontroller vid skanning I var dock fortfarande observerbar i tvärsnittsanalysen vid skanning IV. Den relativa ökningen av grå substans över tiden är därför subtil, dvs inte tillräckligt distinkt för att ha en effekt på tvärsnittsanalysen, ett fynd som redan har visats i studier som undersöker erfarenhetsberoende plasticitet [30], [31]. Vi noterar att det faktum att vi visar att vissa delar av hjärnförändringar på grund av kronisk smärta är reversibla inte utesluter att vissa andra delar av dessa förändringar är irreversibla.
Intressant nog observerade vi att minskningen av den grå substansen i ACC hos patienter med kronisk smärta före operation verkar fortsätta 6 veckor efter operationen (skanning II) och bara ökar mot skanning III och IV, möjligen på grund av smärta efter operationen, eller minskning av motoriken. fungera. Detta är i linje med beteendedata för den fysiska mobilitetspoängen som ingår i NHP, som postoperativt inte visade någon signifikant förändring vid tidpunkt II utan ökade signifikant mot skanning III och IV. Notera att våra patienter inte rapporterade någon smärta i höften efter operationen, men upplevde smärta efter operationen i omgivande muskler och hud som uppfattades väldigt olika av patienterna. Men eftersom patienter fortfarande rapporterade en del smärta vid skanning II, kontrasterade vi också den första skanningen (före operationen) med skanningarna III+IV (efter operationen), vilket avslöjade en ökning av grå substans i frontal cortex och motor cortex. Vi noterar att denna kontrast är mindre sträng på grund av färre skanningar per tillstånd (smärta vs. icke-smärta). När vi sänkte tröskeln upprepar vi vad vi har hittat med kontrast av I+II mot III+IV.
Våra data tyder starkt på att förändringar i grå substans hos patienter med kronisk smärta, som vanligtvis finns i områden som är involverade i supraspinal nociceptiv bearbetning [4] varken beror på neuronal atrofi eller hjärnskada. Det faktum att dessa förändringar som ses i det kroniska smärttillståndet inte vänder helt kan förklaras med den relativt korta observationsperioden (ett år efter operationen kontra i genomsnitt sju år av kronisk smärta före operationen). Neuroplastiska hjärnförändringar som kan ha utvecklats under flera år (som en konsekvens av konstant nociceptiv input) behöver förmodligen mer tid för att vända helt. En annan möjlighet till varför ökningen av grå substans bara kan detekteras i longitudinella data men inte i tvärsnittsdata (dvs mellan kohorter vid tidpunkt IV) är att antalet patienter (n?=?20) är för litet. Det måste påpekas att variansen mellan hjärnor hos flera individer är ganska stor och att longitudinella data har fördelen att variansen är relativt liten då samma hjärnor skannas flera gånger. Följaktligen kommer subtila förändringar endast att kunna detekteras i longitudinella data [30], [31], [32]. Naturligtvis kan vi inte utesluta att dessa förändringar är åtminstone delvis irreversibla även om det är osannolikt, med tanke på resultaten av träningsspecifik strukturell plasticitet och omorganisation [4], [12], [30], [33], [34]. För att svara på denna fråga måste framtida studier undersöka patienter upprepade gånger över längre tidsramar, möjligen år.
Vi noterar att vi bara kan dra begränsade slutsatser om dynamiken i morfologiska hjärnförändringar över tid. Anledningen är att när vi designade den här studien 2007 och scannade 2008 och 2009 var det inte känt om strukturella förändringar skulle inträffa överhuvudtaget och av genomförbarhetsskäl valde vi skanningsdatum och tidsramar som beskrivs här. Man skulle kunna hävda att den grå substansen förändras i tid, som vi beskriver för patientgruppen, kan ha hänt i kontrollgruppen också (tidseffekt). Eventuella förändringar på grund av åldrande, om alls, skulle dock förväntas vara en minskning i volym. Med tanke på vår a priori hypotes, baserad på 9 oberoende studier och kohorter som visar minskningar av grå substans hos patienter med kronisk smärta [7], [8], [9], [15], [24], [25], [26], [27], [28], fokuserade vi på regionala ökningar över tid och tror därför att vårt fynd inte är en enkel tidseffekt. Notera att vi inte kan utesluta att den grå substansen minskar med tiden som vi hittade i vår patientgrupp kan bero på en tidseffekt, eftersom vi inte har skannat vår kontrollgrupp inom samma tidsram. Givet resultaten bör framtida studier syfta till fler och kortare tidsintervall, givet att träningsberoende morfometriska hjärnförändringar kan inträffa så snabbt som efter 1 vecka [32], [33].
Förutom effekten av den nociceptiva aspekten av smärta på hjärnans grå substans [17], [34] observerade vi att förändringar i motorisk funktion förmodligen också bidrar till de strukturella förändringarna. Vi fann att motoriska och premotoriska områden (område 6) ökade över alla tidsintervall (Figur 3). Intuitivt kan detta bero på förbättring av motorisk funktion över tid eftersom patienterna inte längre var begränsade i att leva ett normalt liv. Noterbart fokuserade vi inte på motorisk funktion utan en förbättring av smärtupplevelsen, med tanke på vår ursprungliga strävan att undersöka om den välkända minskningen av hjärnans grå substans hos patienter med kronisk smärta i princip är reversibel. Följaktligen använde vi inte specifika instrument för att undersöka motorisk funktion. Ändå är (funktionell) motorisk cortex-omorganisation hos patienter med smärtsyndrom väl dokumenterad [35], [36], [37], [38]. Dessutom är den motoriska cortex ett mål i terapeutiska tillvägagångssätt hos medicinskt svårlösta patienter med kronisk smärta som använder direkt hjärnstimulering [39], [40], transkraniell likströmsstimulering [41] och repetitiv transkraniell magnetisk stimulering [42], [43]. De exakta mekanismerna för sådan modulering (lättnad vs. hämning, eller helt enkelt störning i de smärtrelaterade nätverken) är ännu inte klarlagda [40]. En nyligen genomförd studie visade att en specifik motorisk upplevelse kan förändra hjärnans struktur [13]. Synaptogenes, omorganisation av rörelserepresentationer och angiogenes i motorisk cortex kan förekomma med speciella krav på en motorisk uppgift. Tsao et al. visade omorganisation i den motoriska cortex hos patienter med kronisk ländryggssmärta som verkar vara ryggsmärtaspecifika [44] och Puri et al. observerade en minskning av vänster kompletterande motorområdes grå substans hos fibromyalgipatienter [45]. Vår studie var inte utformad för att reda ut de olika faktorer som kan förändra hjärnan vid kronisk smärta, men vi tolkar våra data om förändringarna i den grå substansen att de inte enbart speglar konsekvenserna av konstant nociceptiv input. Faktum är att en nyligen genomförd studie av patienter med neuropatisk smärta påpekade abnormiteter i hjärnregioner som omfattar emotionell, autonom och smärtuppfattning, vilket antyder att de spelar en avgörande roll i den globala kliniska bilden av kronisk smärta [28].
Figur 3: Statistiska parametriska kartor som visar en signifikant ökning av hjärnans grå substans i motoriska områden (område 6) hos patienter med coxarthrosis före jämfört med efter THR (longitudinell analys, skanning I Kontrastuppskattningar till x?=?19, y?=??12, z?=?70.
Två nyligen genomförda pilotstudier fokuserade på höftprotesbehandling hos artrospatienter, det enda kroniska smärtsyndromet som huvudsakligen kan botas med total höftprotes [17], [46] och dessa data flankeras av en mycket ny studie på patienter med kronisk ländryggssmärta. 47]. Dessa studier måste ses i ljuset av flera longitudinella studier som undersöker erfarenhetsberoende neuronal plasticitet hos människor på en strukturell nivå [30], [31] och en nyligen genomförd studie om strukturella förändringar i hjärnan hos friska frivilliga som upplever upprepad smärtsam stimulering [34] . Nyckelbudskapet i alla dessa studier är att den huvudsakliga skillnaden i hjärnans struktur mellan smärtpatienter och kontroller kan avta när smärtan är botad. Det måste dock beaktas att det helt enkelt inte är klart om förändringarna hos patienter med kronisk smärta enbart beror på nociceptiv input eller på konsekvenserna av smärta eller både och. Det är mer än troligt att beteendeförändringar, såsom deprivation eller förstärkning av sociala kontakter, smidighet, fysisk träning och livsstilsförändringar är tillräckliga för att forma hjärnan [6], [12], [28], [48]. Särskilt depression som en komorbiditet eller konsekvens av smärta är en nyckelkandidat för att förklara skillnaderna mellan patienter och kontroller. En liten grupp av våra patienter med OA visade milda till måttliga depressiva symtom som förändrades med tiden. Vi fann inte att de strukturella förändringarna samvarierade signifikant med BDI-poängen men frågan uppstår hur många andra beteendeförändringar på grund av frånvaro av smärta och motorisk förbättring som kan bidra till resultaten och i vilken utsträckning de gör det. Dessa beteendeförändringar kan möjligen påverka en minskning av grå substans i kronisk smärta såväl som en ökning av grå substans när smärtan är borta.
En annan viktig faktor som kan påverka vår tolkning av resultaten är det faktum att nästan alla patienter med kronisk smärta tog mediciner mot smärta, som de slutade när de var smärtfria. Man skulle kunna hävda att NSAID som diklofenak eller ibuprofen har vissa effekter på nervsystemet och detsamma gäller för opioider, antiepileptika och antidepressiva, mediciner som ofta används vid kronisk smärtbehandling. Effekten av smärtstillande medel och andra mediciner på morfometriska fynd kan mycket väl vara viktig (48). Ingen studie har hittills visat effekter av smärtstillande medicin på hjärnans morfologi men flera artiklar fann att förändringar i hjärnstrukturen hos patienter med kronisk smärta varken enbart förklaras av smärtrelaterad inaktivitet [15], eller av smärtstillande medicin [7], [9], [49]. Däremot saknas specifika studier. Ytterligare forskning bör fokusera på de erfarenhetsberoende förändringarna i kortikal plasticitet, vilket kan ha stora kliniska implikationer för behandling av kronisk smärta.
Vi fann också minskningar av grå substans i den longitudinella analysen, möjligen på grund av omorganiseringsprocesser som åtföljer förändringar i motorisk funktion och smärtuppfattning. Det finns lite information tillgänglig om longitudinella förändringar i hjärnans grå substans vid smärttillstånd, av denna anledning har vi ingen hypotes för en minskning av grå substans i dessa områden efter operationen. Teutsch et al. [25] fann en ökning av hjärnans grå substans i den somatosensoriska och mellancingulära cortex hos friska frivilliga som upplevde smärtsam stimulering i ett dagligt protokoll under åtta dagar i följd. Upptäckten av ökning av grå substans efter experimentell nociceptiv inmatning överlappade anatomiskt till viss del minskningen av hjärnans grå substans i denna studie hos patienter som botades från långvarig kronisk smärta. Detta innebär att nociceptiv insats hos friska frivilliga leder till träningsberoende strukturella förändringar, som det möjligen gör hos patienter med kronisk smärta, och att dessa förändringar reverserar hos friska frivilliga när nociceptiv inmatning upphör. Följaktligen kunde minskningen av grå substans i dessa områden som ses hos patienter med OA tolkas att följa samma grundläggande process: träningsberoende förändringar hjärnförändringar [50]. Som en icke-invasiv procedur är MR-morfometri det idealiska verktyget för att hitta de morfologiska substraten för sjukdomar, fördjupa vår förståelse av sambandet mellan hjärnans struktur och funktion, och även för att övervaka terapeutiska ingrepp. En av de stora utmaningarna i framtiden är att anpassa detta kraftfulla verktyg för multicenter och terapeutiska prövningar av kronisk smärta.
Begränsningar för denna studie
Även om den här studien är en förlängning av vår tidigare studie som utökar uppföljningsdata till 12 månader och undersöker fler patienter, är vår principiella upptäckt att morfometriska hjärnförändringar vid kronisk smärta är reversibla ganska subtil. Effektstorlekarna är små (se ovan) och effekterna drivs delvis av en ytterligare minskning av den regionala hjärnans grå substans volym vid tidpunkten för skanning 2. När vi exkluderar data från skanning 2 (direkt efter operationen) är endast signifikant ökningar i hjärnans grå substans för motorisk cortex och frontal cortex överlever en tröskel på p<0.001 okorrigerat (tabell 3).
Slutsats
Det är inte möjligt att urskilja i vilken utsträckning de strukturella förändringarna vi observerade beror på förändringar i nociceptiv input, förändringar i motorisk funktion eller läkemedelskonsumtion eller förändringar i välbefinnande som sådant. Att maskera gruppkontrasterna för den första och sista skanningen med varandra visade mycket mindre skillnader än förväntat. Förmodligen utvecklas hjärnförändringar på grund av kronisk smärta med alla konsekvenser under en ganska lång tidsperiod och kan också behöva lite tid för att återgå. Ändå avslöjar dessa resultat omorganisationsprocesser, vilket starkt tyder på att kronisk nociceptiv input och motorisk försämring hos dessa patienter leder till förändrad bearbetning i kortikala regioner och följaktligen strukturella hjärnförändringar som i princip är reversibla.
Erkännanden
Vi tackar alla frivilliga för deltagandet i denna studie och gruppen Fysik och metoder på NeuroImage Nord i Hamburg. Studien fick etiskt godkännande av den lokala etiska kommittén och skriftligt informerat samtycke erhölls från alla studiedeltagare innan undersökningen.
Finansieringsdeklaration
Detta arbete stöddes av anslag från DFG (Tyska forskningsstiftelsen) (MA 1862/2-3) och BMBF (Federala ministeriet för utbildning och forskning) (371 57 01 och NeuroImage Nord). Finansiärerna hade ingen roll i studiedesign, datainsamling och analys, beslut att publicera eller förberedelse av manuskriptet.
Endocannabinoidsystemet: Det väsentliga systemet du aldrig har hört talas om
Om du inte har hört talas om det endocannabinoida systemet, eller ECS, behöver du inte känna dig generad. Tillbaka på 1960-talet isolerade de utredare som blev intresserade av cannabis bioaktivitet så småningom många av dess aktiva kemikalier. Det tog dock ytterligare 30 år för forskare som studerade djurmodeller att hitta en receptor för dessa ECS-kemikalier i hjärnan på gnagare, en upptäckt som öppnade en hel värld av undersökningar om ECS-receptorernas existens och vad deras fysiologiska syfte är.
Vi vet nu att de flesta djur, från fiskar till fåglar till däggdjur, har en endocannabinoid, och vi vet att människor inte bara gör sina egna cannabinoider som interagerar med just detta system, utan vi producerar även andra föreningar som interagerar med ECS, de av som observeras i många olika växter och livsmedel, långt bortom Cannabis-arterna.
Som ett system i människokroppen är ECS inte en isolerad strukturell plattform som nervsystemet eller det kardiovaskulära systemet. Istället är ECS en uppsättning receptorer brett spridda över hela kroppen som aktiveras genom en uppsättning ligander som vi gemensamt känner till som endocannabinoider, eller endogena cannabinoider. Båda verifierade receptorer kallas bara CB1 och CB2, även om det finns andra som föreslogs. PPAR- och TRP-kanaler förmedlar också vissa funktioner. På samma sätt hittar du bara två väldokumenterade endocannabinoider: anadamid och 2-arakidonoylglycerol, eller 2-AG.
Grundläggande för det endocannabinoida systemet är dessutom de enzymer som syntetiserar och bryter ner endocannabinoiderna. Endocannabinoider tros syntetiseras i en grund som behövs. De primära involverade enzymerna är diacylglycerollipas och N-acyl-fosfatidyletanolamin-fosfolipas D, som syntetiserar 2-AG respektive anandamid. De två huvudsakliga nedbrytande enzymerna är fettsyraamidhydrolas, eller FAAH, som bryter ner anandamid, och monoacylglycerollipas, eller MAGL, som bryter ner 2-AG. Regleringen av dessa två enzymer kan öka eller minska moduleringen av ECS.
Vad är ECS:s funktion?
ECS är kroppens huvudsakliga homeostatiska regleringssystem. Det kan lätt ses som kroppens interna adaptogena system, som alltid arbetar för att upprätthålla balansen mellan olika funktioner. Endocannabinoider fungerar i stort sett som neuromodulatorer och som sådana reglerar de ett brett spektrum av kroppsliga processer, från fertilitet till smärta. Några av de mer kända funktionerna från ECS är följande:
Nervsystem
Från det centrala nervsystemet, eller CNS, kommer allmän stimulering av CB1-receptorerna att hämma frisättningen av glutamat och GABA. I CNS spelar ECS en roll i minnesbildning och inlärning, främjar neurogenes i hippocampus, reglerar också neuronal excitabilitet. ECS spelar också en roll i hur hjärnan kommer att reagera på skador och inflammation. Från ryggmärgen modulerar ECS smärtsignalering och ökar naturlig analgesi. I det perifera nervsystemet, där CB2-receptorer kontrollerar, verkar ECS främst i det sympatiska nervsystemet för att reglera funktioner i tarm-, urin- och reproduktionsorganen.
Stress och humör
ECS har flera effekter på stressreaktioner och känslomässig reglering, såsom initiering av detta kroppsliga svar på akut stress och anpassning över tid till mer långsiktiga känslor, såsom rädsla och ångest. Ett hälsosamt fungerande endocannabinoidsystem är avgörande för hur människor modulerar mellan en tillfredsställande grad av upphetsning jämfört med en nivå som är överdriven och obehaglig. ECS spelar också en roll i minnesbildningen och möjligen särskilt i det sätt på vilket hjärnan prickar in minnen från stress eller skada. Eftersom ECS modulerar frisättningen av dopamin, noradrenalin, serotonin och kortisol, kan det också i stor utsträckning påverka känslomässiga svar och beteenden.
Matsmältningssystemet
Matsmältningskanalen är befolkad med både CB1- och CB2-receptorer som reglerar flera viktiga aspekter av GI-hälsa. Man tror att ECS kan vara den "saknade länken" för att beskriva den tarm-hjärna-immuna länken som spelar en betydande roll för den funktionella hälsan i matsmältningskanalen. ECS är en regulator av tarmimmunitet, kanske genom att begränsa immunsystemet från att förstöra frisk flora, och även genom modulering av cytokinsignalering. ECS modulerar det naturliga inflammatoriska svaret i matsmältningskanalen, vilket har viktiga konsekvenser för ett brett spektrum av hälsoproblem. Gastrisk och allmän gastrointestinal motilitet verkar också delvis styras av ECS.
Aptit och metabolism
ECS, särskilt CB1-receptorerna, spelar en roll i aptit, metabolism och reglering av kroppsfett. Stimulering av CB1-receptorerna höjer det matsökande beteendet, ökar medvetenheten om lukt, reglerar också energibalansen. Både djur och människor som är överviktiga har ECS dysreglering som kan leda till att detta system blir hyperaktivt, vilket bidrar till både överätande och minskad energiförbrukning. Cirkulerande nivåer av anandamid och 2-AG har visat sig vara förhöjda vid fetma, vilket delvis kan bero på minskad produktion av det FAAH-nedbrytande enzymet.
Immunhälsa och inflammatoriskt svar
Immunsystemets celler och organ är rika på endocannabinoidreceptorer. Cannabinoidreceptorer uttrycks i tymuskörteln, mjälten, tonsiller och benmärg, såväl som på T- och B-lymfocyter, makrofager, mastceller, neutrofiler och naturliga mördarceller. ECS anses vara den primära drivkraften för immunsystemets balans och homeostas. Även om inte alla funktioner hos ECS från immunsystemet förstås, verkar ECS reglera cytokinproduktionen och även ha en roll i att förhindra överaktivitet i immunsystemet. Inflammation är en naturlig del av immunsvaret, och den spelar en mycket normal roll vid akuta förolämpningar mot kroppen, inklusive skador och sjukdomar; men när den inte hålls i schack kan den bli kronisk och bidra till en kaskad av negativa hälsoproblem, såsom kronisk smärta. Genom att hålla immunsvaret i schack hjälper ECS till att upprätthålla ett mer balanserat inflammatoriskt svar genom kroppen.
Andra hälsoområden som regleras av ECS:
Benhälsa
Fertilitet
hudens hälsa
Arteriell och respiratorisk hälsa
Sömn och dygnsrytm
Hur man bäst stödjer ett hälsosamt ECS är en fråga som många forskare nu försöker svara på. Håll utkik för mer information om detta framväxande ämne.
SammanfattningsvisKronisk smärta har associerats med hjärnförändringar, inklusive minskning av grå substans. Men artikeln ovan visade att kronisk smärta kan förändra hjärnans övergripande struktur och funktion. Även om kronisk smärta kan leda till dessa, bland andra hälsoproblem, kan korrekt behandling av patientens underliggande symtom vända förändringar i hjärnan och reglera grå substans. Dessutom har fler och fler forskningsstudier dykt upp bakom vikten av det endocannabinoida systemet och dess funktion för att kontrollera och hantera kronisk smärta och andra hälsoproblem. Information som refereras från National Center for Biotechnology Information (NCBI).�Omfattningen av vår information är begränsad till kiropraktik såväl som till ryggradsskador och tillstånd. För att diskutera ämnet är du välkommen att fråga Dr Jimenez eller kontakta oss på�915-850-0900 .
Curated av Dr. Alex Jimenez
Ytterligare ämnen: Ryggsmärta
Ryggont är en av de vanligaste orsakerna till funktionshinder och missade dagar på jobbet över hela världen. Faktum är att ryggsmärta har tillskrivits som den näst vanligaste anledningen till doktorsbesök, som endast överträffas av övre luftvägsinfektioner. Cirka 80 procent av befolkningen kommer att uppleva någon typ av ryggsmärta minst en gång i livet. Ryggraden är en komplex struktur bestående av ben, leder, ligament och muskler, bland annat mjuka vävnader. På grund av detta, skador och / eller förvärrade förhållanden, till exempel hernierade skivor, kan så småningom leda till symtom på ryggont. Sportskador eller skador på bilolyckor är ofta den vanligaste orsaken till ryggsmärta, men ibland kan de enklaste rörelserna få ont. Lyckligtvis kan alternativa behandlingsalternativ, som kiropraktikvård, hjälpa till att lindra ryggont genom användningen av ryggmärgsjusteringar och manuell manipulation, vilket i slutändan förbättrar smärtlindringen.
2.�Flor H, Nikolajsen L, Staehelin Jensen T (2006)�Fantomsmärta i extremiteterna: ett fall av maladaptiv CNS-plasticitet?�Nat Rev Neurosci�7: 873�881.�[PubMed]
3.�Wrigley PJ, Gustin SM, Macey PM, Nash PG, Gandevia SC, et al. (2009)�Anatomiska förändringar i mänskliga motoriska cortex och motoriska vägar efter fullständig thorax ryggmärgsskada. Cereb Cortex�19: 224�232.�[PubMed]
4.�maj A (2008)�Kronisk smärta kan förändra hjärnans struktur. Smärta�137: 7�15.�[PubMed]
5.�May A (2009) Morphing voxels: hypen kring strukturell avbildning av huvudvärkpatienter. Hjärna.[PubMed]
6.�Apkarian AV, Baliki MN, Geha PY (2009)�Mot en teori om kronisk smärta. Prog Neurobiol�87: 81�97.�[PMC gratis artikel]�[PubMed]
7.�Apkarian AV, Sosa Y, Sonty S, Levy RM, Harden RN, et al. (2004)�Kronisk ryggsmärta är associerad med minskad prefrontal och thalamus grå substans densitet. J Neurosci�24: 10410�10415.�[PubMed]
8.�Rocca MA, Ceccarelli A, Falini A, Colombo B, Tortorella P, et al. (2006)�Förändringar i hjärnans grå substans hos migränpatienter med T2-synliga lesioner: en 3-T MRI-studie. Stroke�37: 1765�1770.�[PubMed]
9.�Kuchinad A, Schweinhardt P, Seminowicz DA, Wood PB, Chizh BA, et al. (2007)�Accelererad förlust av grå substans i hjärnan hos fibromyalgipatienter: för tidigt åldrande av hjärnan?�J Neurosci�27: 4004–4007.[PubMed]
10.�Tracey I, Bushnell MC (2009)�Hur neuroimaging studier har utmanat oss att tänka om: är kronisk smärta en sjukdom?�J Smärta�10: 1113�1120.�[PubMed]
11.�Franke K, Ziegler G, Kloppel S, Gaser C (2010)�Uppskattning av åldern på friska försökspersoner från T1-viktade MRI-skanningar med hjälp av kärnmetoder: utforska påverkan av olika parametrar. NeuroImage�50: 883�892.�[PubMed]
12.�Draganski B, May A (2008)�Träningsinducerade strukturella förändringar i den vuxna mänskliga hjärnan. Behav Brain Res�192: 137�142.�[PubMed]
13.�Adkins DL, Boychuk J, Remple MS, Kleim JA (2006)�Motorisk träning inducerar erfarenhetsspecifika mönster av plasticitet över motorisk cortex och ryggmärg. J Appl Physiol�101: 1776�1782.�[PubMed]
14.�Duerden EG, Laverdure-Dupont D (2008)�Övning gör cortex. J Neurosci�28: 8655�8657.�[PubMed]
15.�Draganski B, Moser T, Lummel N, Ganssbauer S, Bogdahn U, et al. (2006)�Minskning av thalamisk grå substans efter amputation av extremiteter. NeuroImage�31: 951�957.�[PubMed]
16.�Nikolajsen L, Brandsborg B, Lucht U, Jensen TS, Kehlet H (2006)�Kronisk smärta efter total höftprotesplastik: en rikstäckande frågeformulärstudie. Acta Anaesthesiol Scand�50: 495�500.�[PubMed]
17.�Rodriguez-Raecke R, Niemeier A, Ihle K, Ruether W, May A (2009)�Hjärnans grå substans minskning av kronisk smärta är konsekvensen och inte orsaken till smärta. J Neurosci�29: 13746�13750.�[PubMed]
18.�Beck AT, Ward CH, Mendelson M, Mock J, Erbaugh J (1961)�En inventering för att mäta depression. Arch Gen Psychiatry�4: 561�571.�[PubMed]
19.�Franke G (2002) Symptom-Checklista nach LR Derogatis – Manual. G�ttingen Beltz Test Verlag.
20.�Geissner E (1995) The Pain Perception Scale - en differentierad och förändringskänslig skala för att bedöma kronisk och akut smärta. Rehabilitering (Stuttg) 34: XXXV�XLIII.�[PubMed]
21.�Bullinger M, Kirchberger I (1998) SF-36 – Fragebogen zum Gesundheitszustand. Hand-anweisung. Göttingen: Hogrefe.
23.�Bra CD, Johnsrude IS, Ashburner J, Henson RN, Friston KJ, et al. (2001)�En voxelbaserad morfometrisk studie av åldrande i 465 normala vuxna mänskliga hjärnor. NeuroImage�14: 21�36.�[PubMed]
24.�Baliki MN, Chialvo DR, Geha PY, Levy RM, Harden RN, et al. (2006)�Kronisk smärta och den känslomässiga hjärnan: specifik hjärnaktivitet associerad med spontana fluktuationer i intensiteten av kronisk ryggsmärta. J Neurosci�26: 12165�12173.�[PMC gratis artikel]�[PubMed]
25.�Lutz J, Jager L, de Quervain D, Krauseneck T, Padberg F, et al. (2008)�Avvikelser i vita och grå substanser i hjärnan hos patienter med fibromyalgi: en diffusionstensor- och volymetrisk avbildningsstudie. Artrit Rheum�58: 3960�3969.�[PubMed]
26.�Wrigley PJ, Gustin SM, Macey PM, Nash PG, Gandevia SC, et al. (2008)�Anatomiska förändringar i mänskliga motoriska cortex och motoriska banor efter komplett thorax ryggmärgsskada. Cereb Cortex19: 224�232.�[PubMed]
27.�Schmidt-Wilcke T, Hierlmeier S, Leinisch E (2010) Förändrad regional hjärnmorfologi hos patienter med kronisk ansiktssmärta. Huvudvärk.�[PubMed]
28.�Geha PY, Baliki MN, Harden RN, Bauer WR, Parrish TB, et al. (2008)�Hjärnan vid kronisk CRPS-smärta: onormala interaktioner med grå-vit substans i känslomässiga och autonoma regioner. Neuron�60: 570�581.�[PMC gratis artikel]�[PubMed]
29.�Brazier J, Roberts J, Deverill M (2002)�Uppskattningen av ett preferensbaserat hälsomått från SF-36. J Health Econ�21: 271�292.�[PubMed]
30.�Draganski B, Gaser C, Busch V, Schuierer G, Bogdahn U, et al. (2004)�Neuroplasticitet: förändringar i grå substans inducerad genom träning. Natur�427: 311�312.�[PubMed]
31.�Boyke J, Driemeyer J, Gaser C, Buchel C, May A (2008)�Utbildningsinducerad hjärnstruktur förändras hos äldre. J Neurosci�28: 7031�7035.�[PubMed]
32.�Driemeyer J, Boyke J, Gaser C, Buchel C, May A (2008)�Förändringar i grå substans framkallade av lärande återbesöks. PLoS ONE�3: e2669.�[PMC gratis artikel]�[PubMed]
33.�May A, Hajak G, Ganssbauer S, Steffens T, Langguth B, et al. (2007)�Strukturella hjärnförändringar efter 5 dagars intervention: dynamiska aspekter av neuroplasticitet. Cereb Cortex�17: 205�210.�[PubMed]
34.�Teutsch S, Herken W, Bingel U, Schoell E, May A (2008)�Förändringar i hjärnans grå substans på grund av upprepad smärtsam stimulering. NeuroImage�42: 845�849.�[PubMed]
35.�Flor H, Braun C, Elbert T, Birbaumer N (1997)�Omfattande omorganisation av primär somatosensorisk cortex hos patienter med kronisk ryggsmärta. Neurosci Lett�224: 5�8.�[PubMed]
36.�Flor H, Denke C, Schaefer M, Grusser S (2001)�Effekt av sensorisk diskrimineringsträning på kortikal omorganisation och fantomsmärta i extremiteterna. Lansetten�357: 1763�1764.�[PubMed]
37.�Swart CM, Stins JF, Beek PJ (2009)�Kortikala förändringar i komplext regionalt smärtsyndrom (CRPS). Eur J Pain�13: 902�907.�[PubMed]
38.�Maihofner C, Baron R, DeCol R, Binder A, Birklein F, et al. (2007)�Det motoriska systemet visar adaptiva förändringar i komplext regionalt smärtsyndrom. Hjärna�130: 2671�2687.�[PubMed]
39.�Fontaine D, Hamani C, Lozano A (2009)�Effekt och säkerhet av motorisk cortexstimulering för kronisk neuropatisk smärta: kritisk granskning av litteraturen. J Neurosurg�110: 251�256.�[PubMed]
40.�Levy R, Deer TR, Henderson J (2010)�Intrakraniell neurostimulering för smärtkontroll: en översyn. Smärtläkare�13: 157�165.�[PubMed]
41.�Antal A, Brepohl N, Poreisz C, Boros K, Csifcsak G, et al. (2008)�Transkraniell likströmsstimulering över somatosensorisk cortex minskar experimentellt inducerad akut smärtuppfattning. Clin J Smärta24: 56�63.�[PubMed]
42.�Teepker M, Hotzel J, Timmesfeld N, Reis J, Mylius V, et al. (2010)�Lågfrekvent rTMS av vertex vid profylaktisk behandling av migrän. Cephalalgia�30: 137�144.�[PubMed]
43.�O�Connell N, Wand B, Marston L, Spencer S, Desouza L (2010)�Icke-invasiva tekniker för hjärnstimulering för kronisk smärta. En rapport om en Cochrane systematisk granskning och metaanalys. Eur J Phys Rehabil Med�47: 309�326.�[PubMed]
44.�Tsao H, Galea MP, Hodges PW (2008)�Omorganisering av den motoriska cortex är förknippad med postural kontrollbrist vid återkommande ländryggssmärtor. Hjärna�131: 2161�2171.�[PubMed]
45.�Puri BK, Agour M, Gunatilake KD, Fernando KA, Gurusinghe AI, et al. (2010)�Minskning av grå substans i vänster kompletterande motorområde hos vuxna kvinnliga fibromyalgipatienter med markant trötthet och utan affektiv störning: en pilotkontrollerad 3-T magnetisk resonanstomografi voxelbaserad morfometristudie. J Int Med Res�38: 1468�1472.�[PubMed]
46.�Gwilym SE, Fillipini N, Douaud G, Carr AJ, Tracey I (2010) Talamusatrofi associerad med smärtsam artros i höften är reversibel efter artroplastik; en longitudinell voxelbaserad morfometrisk studie. Artrit Rheum.�[PubMed]
47.�Seminowicz DA, Wideman TH, Naso L, Hatami-Khoroushahi Z, Fallatah S, et al. (2011)�Effektiv behandling av kronisk ländryggssmärta hos människor vänder onormal hjärnans anatomi och funktion. J Neurosci31: 7540�7550.�[PubMed]
48.�May A, Gaser C (2006)�Magnetisk resonansbaserad morfometri: ett fönster till hjärnans strukturella plasticitet. Curr Opin Neurol�19: 407�411.�[PubMed]
49.�Schmidt-Wilcke T, Leinisch E, Straube A, Kampfe N, Draganski B, et al. (2005)�Grå substans minskning hos patienter med kronisk huvudvärk av spänningstyp. Neurologi�65: 1483�1486.�[PubMed]
50.�maj A (2009)�Morphing voxels: hypen kring strukturell bildbehandling av huvudvärkpatienter. Hjärna 132(Pt6): 1419�1425.�[PubMed]
Smärtans biokemi:�Alla smärtsyndrom har en inflammationsprofil. En inflammatorisk profil kan variera från person till person och kan även variera hos en person vid olika tidpunkter. Behandlingen av smärtsyndrom är att förstå denna inflammationsprofil. Smärtsyndrom behandlas medicinskt, kirurgiskt eller båda. Målet är att hämma/hämma produktionen av inflammatoriska mediatorer. Och ett framgångsrikt resultat är ett som resulterar i mindre inflammation och naturligtvis mindre smärta.
Biokemi av smärta
mål:
Vilka är nyckelspelarna
Vilka är de biokemiska mekanismerna?
Vilka är konsekvenserna?
Inflammationsrecension:
Nyckelspelare
Varför gör min axel ont? En genomgång av den neuroanatomiska och biokemiska grunden för axelsmärta
SAMMANDRAG
Om en patient frågar �varför gör min axel ont?� kommer samtalet snabbt att övergå till vetenskaplig teori och ibland ogrundade gissningar. Klinikern blir ofta medveten om gränserna för den vetenskapliga grunden för deras förklaring, vilket visar på ofullständigheten i vår förståelse av axelsmärtans natur. Denna recension tar ett systematiskt tillvägagångssätt för att hjälpa till att besvara grundläggande frågor om axelsmärta, i syfte att ge insikter i framtida forskning och nya metoder för behandling av axelsmärta. Vi ska utforska rollerna för (1) de perifera receptorerna, (2) perifer smärtbehandling eller �nociception�, (3) ryggmärgen, (4) hjärnan, (5) receptorernas placering i axeln och (6) ) axelns neurala anatomi. Vi överväger också hur dessa faktorer kan bidra till variationen i den kliniska presentationen, diagnosen och behandlingen av axelsmärtor. På detta sätt syftar vi till att ge en överblick över beståndsdelarna i det perifera smärtdetekteringssystemet och centrala smärtbearbetningsmekanismer vid axelsmärta som samverkar för att ge klinisk smärta.
INTRODUKTION: EN MYCKET KORT HISTORIA OM SMÄRTVETENSKAP VIKTIGT FÖR KLINIKER
Smärtans natur i allmänhet har varit föremål för mycket kontroverser under det senaste århundradet. På 17-talet föreslog Descartes teori1 att smärtans intensitet var direkt relaterad till mängden associerad vävnadsskada och att smärta bearbetades i en distinkt väg. Många tidigare teorier förlitade sig på denna så kallade "dualistiska" Descartianska filosofi, och såg smärta som en konsekvens av stimuleringen av en "specifik" perifer smärtreceptor i hjärnan. På 20-talet uppstod en vetenskaplig kamp mellan två motsatta teorier, nämligen specificitetsteori och mönsterteori. Den Descartianska �specificitetsteorin� såg smärta som en specifik separat modalitet för sensorisk inmatning med sin egen apparat, medan �mönsterteorin� ansåg att smärta var ett resultat av intensiv stimulering av ospecifika receptorer.2 1965, Wall och Melzacks 3 grinteori om smärta gav bevis för en modell där smärtuppfattningen modulerades av både sensorisk feedback och det centrala nervsystemet. Ett annat stort framsteg inom smärtteorin såg vid ungefär samma tidpunkt upptäckten av opioidernas specifika verkningssätt.4 Senare har de senaste framstegen inom neuroimaging och molekylär medicin avsevärt utökat vår övergripande förståelse av smärta.
Så hur hänger detta ihop med axelvärk?�Axelvärk är ett vanligt kliniskt problem, och en gedigen förståelse för hur smärta bearbetas av kroppen är avgörande för att på bästa sätt kunna diagnostisera och behandla en patients smärta. Framsteg i vår kunskap om smärtbehandling lovar att förklara obalansen mellan patologi och uppfattningen av smärta, de kan också hjälpa oss att förklara varför vissa patienter misslyckas med att svara på vissa behandlingar.
GRUNDLÄGGANDE BYGGSTETS AV SMÄRTA
Perifera sensoriska receptorer: mekanoreceptorn och �nociceptorn�
Det finns många typer av perifera sensoriska receptorer närvarande i det mänskliga muskuloskeletala systemet. 5 De kan klassificeras baserat på deras funktion (som mekanoreceptorer, termoreceptorer eller nociceptorer) eller morfologi (fria nervändar eller olika typer av inkapslade receptorer).5 De olika typerna av receptorer kan sedan subklassificeras ytterligare baserat på förekomsten av vissa kemiska markörer. Det finns betydande överlappningar mellan olika funktionella klasser av receptorer, till exempel
Perifer smärtbehandling: �Nociception�
Vävnadsskada involverar en mängd olika inflammatoriska mediatorer som frisätts av skadade celler inklusive bradykinin, histamin, 5-hydroxitryptamin, ATP, kväveoxid och vissa joner (K+ och H+). Aktiveringen av arakidonsyravägen leder till produktion av prostaglandiner, tromboxaner och leukotriener. Cytokiner, inklusive interleukinerna och tumörnekrosfaktorn ?, och neurotrofiner, såsom nervtillväxtfaktor (NGF), frisätts också och är intimt involverade i att underlätta inflammation.15 Andra ämnen som excitatoriska aminosyror (glutamat) och opioider ( endotelin-1) har också varit inblandade i det akuta inflammatoriska svaret.16 17 Vissa av dessa medel kan direkt aktivera nociceptorer, medan andra orsakar rekryteringen av andra celler som sedan frisätter ytterligare underlättande medel.18 Denna lokala process som resulterar i ökad reaktionsförmåga av nociceptiva neuroner till deras normala input och/eller rekryteringen av ett svar på normalt undertröskelvärden kallas �perifer sensibilisering�.�Figur 1 sammanfattar några av de involverade nyckelmekanismerna.
NGF och den transienta receptorpotentiala katjonkanalunderfamiljen V medlem 1 (TRPV1)-receptorn har ett symbiotiskt samband när det kommer till inflammation och nociceptorsensibilisering. Cytokinerna som produceras i inflammerad vävnad resulterar i en ökning av NGF-produktionen.19 NGF stimulerar frisättningen av histamin och serotonin (5-HT3) av mastceller, och sensibiliserar även nociceptorer, vilket möjligen förändrar egenskaperna hos A? fibrer så att en större andel blir nociceptiva. TRPV1-receptorn finns i en subpopulation av primära afferenta fibrer och aktiveras av capsaicin, värme och protoner. TRPV1-receptorn syntetiseras i den afferenta fiberns cellkropp och transporteras till både de perifera och centrala terminalerna, där den bidrar till känsligheten hos nociceptiva afferenter. Inflammation resulterar i NGF-produktion perifert som sedan binder till tyrosinkinasreceptor typ 1-receptorn på nociceptorterminalerna, NGF transporteras sedan till cellkroppen där det leder till en uppreglering av TRPV1-transkription och följaktligen ökad nociceptorkänslighet.19 20 NGF och andra inflammatoriska mediatorer sensibiliserar också TRPV1 genom en mångfald av sekundära budbärarvägar. Många andra receptorer inklusive kolinerga receptorer, a-aminosmörsyra (GABA)-receptorer och somatostatinreceptorer tros också vara involverade i perifer nociceptorkänslighet.
Ett stort antal inflammatoriska mediatorer har varit specifikt inblandade i axelsmärta och rotatorcuffsjukdom.21�25 Medan vissa kemiska mediatorer direkt aktiverar nociceptorer, leder de flesta till förändringar i själva sensoriska neuronen snarare än att direkt aktivera den. Dessa förändringar kan vara tidig post-translationell eller fördröjd transkription beroende. Exempel på de förstnämnda är förändringar i TRPV1-receptorn eller i spänningsstyrda jonkanaler till följd av fosforylering av membranbundna proteiner. Exempel på det senare inkluderar den NGF-inducerade ökningen av TRV1-kanalproduktion och den kalciuminducerade aktiveringen av intracellulära transkriptionsfaktorer.
Molekylära mekanismer för nociception
Smärtkänslan varnar oss för verklig eller överhängande skada och utlöser lämpliga skyddsreaktioner. Tyvärr överlever smärta ofta sin användbarhet som ett varningssystem och blir istället kronisk och försvagande. Denna övergång till en kronisk fas involverar förändringar i ryggmärgen och hjärnan, men det finns också en anmärkningsvärd modulering där smärtmeddelanden initieras � på nivån av den primära sensoriska neuronen. Ansträngningar för att fastställa hur dessa neuroner upptäcker smärtproducerande stimuli av termisk, mekanisk eller kemisk natur har avslöjat nya signalmekanismer och fört oss närmare att förstå de molekylära händelser som underlättar övergångar från akut till ihållande smärta.
Neurokemin hos nociceptorer
Glutamat är den dominerande excitatoriska signalsubstansen i alla nociceptorer. Histokemiska studier av vuxen DRG avslöjar dock två breda klasser av omyelinerad C-fiber.
Kemiska givare för att göra smärtan värre
Som beskrivits ovan ökar skada vår smärtupplevelse genom att öka nociceptorernas känslighet för både termiska och mekaniska stimuli. Detta fenomen beror delvis på produktion och frisättning av kemiska mediatorer från den primära sensoriska terminalen och från icke-neurala celler (till exempel fibroblaster, mastceller, neutrofiler och blodplättar) i miljön36 (Fig. 3). Vissa komponenter i den inflammatoriska soppan (till exempel protoner, ATP, serotonin eller lipider) kan förändra neuronal excitabilitet direkt genom att interagera med jonkanaler på nociceptorytan, medan andra (till exempel bradykinin och NGF) binder till metabotropa receptorer och förmedla deras effekter genom andra budbärares signalkaskader11. Avsevärda framsteg har gjorts för att förstå den biokemiska grunden för sådana modulerande mekanismer.
Extracellulära protoner och vävnadsacidos
Lokal vävnadsacidos är ett kännetecknande fysiologiskt svar på skada, och graden av associerad smärta eller obehag är väl korrelerad med omfattningen av försurningen37. Applicering av syra (pH 5) på huden producerar ihållande flytningar i en tredjedel eller fler av polymodala nociceptorer som innerverar det receptiva fältet 20.
Cellulära och molekylära smärtmekanismer
Abstrakt
Nervsystemet upptäcker och tolkar ett brett utbud av termiska och mekaniska stimuli samt miljö- och endogena kemiska irriterande ämnen. När de är intensiva genererar dessa stimuli akut smärta, och i samband med ihållande skada uppvisar både perifera och centrala nervsystemets komponenter i smärtöverföringsvägen en enorm plasticitet, vilket förstärker smärtsignaler och producerar överkänslighet. När plasticitet underlättar skyddsreflexer kan det vara fördelaktigt, men när förändringarna kvarstår kan ett kroniskt smärttillstånd uppstå. Genetiska, elektrofysiologiska och farmakologiska studier belyser de molekylära mekanismerna som ligger till grund för upptäckt, kodning och modulering av skadliga stimuli som genererar smärta.
Introduktion: Akut kontra ihållande smärta
Figur 5. Sensibilisering av ryggmärg (central).
Glutamat/NMDA-receptormedierad sensibilisering.� Efter intensiv stimulering eller ihållande skada, aktiverad C och A? nociceptorer frisätter en mängd olika neurotransmittorer inklusive dlutamat, substans P, kalcitoningenrelaterad peptid (CGRP) och ATP, på utgående neuroner i lamina I i det ytliga dorsala hornet (rött). Som en konsekvens kan normalt tysta NMDA-glutamatreceptorer som finns i den postsynaptiska neuronen nu signalera, öka intracellulärt kalcium och aktivera en mängd kalciumberoende signalvägar och andra budbärare inklusive mitogenaktiverat proteinkinas (MAPK), proteinkinas C (PKC) proteinkinas A (PKA) och Src. Denna kaskad av händelser kommer att öka excitabiliteten hos utgångsneuronen och underlätta överföringen av smärtmeddelanden till hjärnan.
Disinhibition.�Under normala omständigheter frisätter hämmande interneuroner (blå) kontinuerligt GABA och/eller glycin (Gly) för att minska excitabiliteten hos lamina I-utgångsneuroner och modulera smärtöverföring (hämmande tonus). Men i samband med skada kan denna hämning gå förlorad, vilket resulterar i hyperalgesi. Dessutom kan disinhibition möjliggöra icke-nociceptiv myeliniserad A? primära afferenter för att koppla in smärtöverföringskretsen så att normalt ofarliga stimuli nu uppfattas som smärtsamma. Detta sker delvis genom desinhiberingen av excitatorisk PKC? uttrycker interneuroner i inre lamina II.
Mikroglial aktivering.�Perifer nervskada främjar frisättning av ATP och kemokin fraktalkin som kommer att stimulera mikrogliaceller. Speciellt resulterar aktivering av purinerga, CX3CR1 och Toll-liknande receptorer på mikroglia (lila) i frisättningen av hjärnhärledd neurotrofisk faktor (BDNF), som genom aktivering av TrkB-receptorer uttryckta av lamina I-utgångsneuroner främjar ökad excitabilitet och förstärkt smärta som svar på både skadlig och ofarlig stimulering (det vill säga hyperalgesi och allodyni). Aktiverade mikroglia frisätter också en mängd cytokiner, såsom tumörnekrosfaktor ? (TNF?), interleukin-1? och 6 (IL-1?, IL-6) och andra faktorer som bidrar till central sensibilisering.
Inflammationens kemiska miljö
Perifer sensibilisering är vanligare ett resultat av inflammationsrelaterade förändringar i den kemiska miljön i nervfibern (McMahon et al., 2008). Sålunda åtföljs vävnadsskada ofta av ackumulering av endogena faktorer som frigörs från aktiverade nociceptorer eller icke-neurala celler som finns inom eller infiltrerar i det skadade området (inklusive mastceller, basofiler, blodplättar, makrofager, neutrofiler, endotelceller, keratinocyter och fibroblaster). Kollektivt. dessa faktorer, som kallas den inflammatoriska soppan, representerar ett brett spektrum av signalmolekyler, inklusive signalsubstanser, peptider (substans P, CGRP, bradykinin), eikosinoider och relaterade lipider (prostaglandiner, tromboxaner, leukotriener, endocannabinoider, cytokiner, neuroestrofiner, cytokiner). och kemokiner, såväl som extracellulära proteaser och protoner. Anmärkningsvärt nog uttrycker nociceptorer en eller flera cellytreceptorer som kan känna igen och svara på var och en av dessa pro-inflammatoriska eller pro-algetiska medel (Figur 4). Sådana interaktioner ökar nervfiberns excitabilitet och ökar därmed dess känslighet för temperatur eller beröring.
Utan tvekan innebär det vanligaste tillvägagångssättet för att minska inflammatorisk smärta att hämma syntesen eller ackumuleringen av komponenter i den inflammatoriska soppan. Detta exemplifieras bäst av icke-steroida antiinflammatoriska läkemedel, såsom aspirin eller ibuprofen, som minskar inflammatorisk smärta och hyperalgesi genom att hämma cyklooxygenaser (Cox-1 och Cox-2) som är involverade i prostaglandinsyntes. Ett andra tillvägagångssätt är att blockera verkan av inflammatoriska medel vid nociceptorn. Här lyfter vi fram exempel som ger ny insikt i cellulära mekanismer för perifer sensibilisering, eller som ligger till grund för nya terapeutiska strategier för behandling av inflammatorisk smärta.
NGF är kanske mest känd för sin roll som en neurotrofisk faktor som krävs för överlevnad och utveckling av sensoriska neuroner under embryogenes, men hos vuxna produceras NGF även vid vävnadsskada och utgör en viktig komponent i den inflammatoriska soppan (Ritner et al. al., 2009). Bland dess många cellulära mål verkar NGF direkt på peptiderga C-fibernociceptorer, som uttrycker NGF-receptortyrosinkinas med hög affinitet, TrkA, såväl som neurotrofinreceptorn med låg affinitet, p75 (Chao, 2003; Snider och McMahon, 1998). NGF producerar djup överkänslighet mot värme och mekaniska stimuli genom två tidsmässigt distinkta mekanismer. Till en början aktiverar en NGF-TrkA-interaktion nedströms signalvägar, inklusive fosfolipas C (PLC), mitogenaktiverat proteinkinas (MAPK) och fosfoinositid 3-kinas (PI3K). Detta resulterar i funktionell potentiering av målproteiner vid den perifera nociceptorterminalen, framför allt TRPV1, vilket leder till en snabb förändring i cellulär och beteendemässig värmekänslighet (Chuang et al., 2001).
Oavsett deras pro-nociceptiva mekanismer har störning av neurotrofin- eller cytokinsignalering blivit en viktig strategi för att kontrollera inflammatorisk sjukdom eller resulterande smärta. Det huvudsakliga tillvägagångssättet involverar blockering av NGF eller TNF-? verkan med en neutraliserande antikropp. När det gäller TNF-a har detta varit anmärkningsvärt effektivt vid behandling av många autoimmuna sjukdomar, inklusive reumatoid artrit, vilket leder till dramatisk minskning av både vävnadsdestruktion och åtföljande hyperalgesi (Atzeni et al., 2005). Eftersom huvudverkan av NGF på den vuxna nociceptorn inträffar vid inflammation, är fördelen med detta tillvägagångssätt att hyperalgesi kommer att minska utan att påverka normal smärtuppfattning. Faktum är att anti-NGF-antikroppar för närvarande är i kliniska prövningar för behandling av inflammatoriska smärtsyndrom (Hefti et al., 2006).
Glutamat/NMDA-receptormedierad sensibilisering
Akut smärta signaleras genom frisättning av glutamat från de centrala terminalerna av nociceptorer, vilket genererar excitatoriska postsynaptiska strömmar (EPSC) i andra ordningens dorsalhornsneuroner. Detta sker främst genom aktivering av postsynaptiska AMPA- och kainatsubtyper av jonotropa glutamatreceptorer. Summering av subtröskel-EPSCs i den postsynaptiska neuronen kommer så småningom att resultera i aktionspotentialavfyrning och överföring av smärtmeddelandet till neuroner av högre ordning.
Andra studier tyder på att förändringar i projektionsneuronen i sig bidrar till den inhiberande processen. Till exempel nedreglerar perifer nervskada djupt K+- Cl-co-transportören KCC2, vilket är avgörande för att upprätthålla normala K+ och Cl- gradienter över plasmamembranet (Coull et al., 2003). Nedreglering av KCC2, som uttrycks i lamina I-projektionsneuroner, resulterar i en förskjutning i Cl-gradienten, så att aktivering av GABA-A-receptorer depolariserar snarare än hyperpolariserar lamina I-projektionsneuronerna. Detta skulle i sin tur förbättra excitabiliteten och öka smärtöverföringen. Faktum är att farmakologisk blockad eller siRNA-medierad nedreglering av KCC2 hos råtta inducerar mekanisk allodyni.
Dela Ebook
Källor:
Varför gör min axel ont? En genomgång av den neuroanatomiska och biokemiska grunden för axelsmärta
Benjamin John Floyd Dean, Stephen Edward Gwilym, Andrew Jonathan Carr
Cellulära och molekylära smärtmekanismer
Allan I. Basbaum1, Diana M. Bautista2, Gregory Scherrer1 och David Julius3
1Department of Anatomy, University of California, San Francisco 94158
2Department of Molecular and Cell Biology, University of California, Berkeley CA 94720 3Department of Physiology, University of California, San Francisco 94158
Molekylära mekanismer för nociception
David Julius* & Allan I. Basbaum�
*Institutionen för cellulär och molekylär farmakologi, och �Department of Anatomy and Physiology och WM Keck Foundation Center for Integrative Neuroscience, University of California San Francisco, San Francisco, Kalifornien 94143, USA (e-post: julius@socrates.ucsf.edu)
Neurogen inflammation, eller NI, är den fysiologiska process där mediatorer släpps ut direkt från de kutana nerverna för att påbörja ett inflammatoriskt svar. Detta resulterar i skapandet av lokala inflammatoriska reaktioner inklusive erytem, svullnad, temperaturökning, ömhet och smärta. Fina omyeliniserade afferenta somatiska C-fibrer, som svarar på lågintensiva mekaniska och kemiska stimuleringar, är till stor del ansvariga för frisättningen av dessa inflammatoriska mediatorer.
När de stimuleras frigör dessa nervbanor i hudnerverna energiska neuropeptider, eller substans P och kalcitoningenrelaterad peptid (CGRP), snabbt in i mikromiljön, vilket utlöser en serie inflammatoriska reaktioner. Det finns en betydande skillnad i immunogen inflammation, det är det allra första skyddande och reparerande svaret som görs av immunsystemet när en patogen kommer in i kroppen, medan neurogen inflammation involverar en direkt koppling mellan nervsystemet och de inflammatoriska svaren. Även om neurogen inflammation och immunologisk inflammation kan existera samtidigt, är de två inte kliniskt oskiljbara. Syftet med artikeln nedan är att diskutera mekanismen för neurogen inflammation och det perifera nervsystemets roll i värdförsvar och immunpatologi.
Neurogen inflammation � Det perifera nervsystemets roll i värdförsvar och immunopatologi
Abstrakt
Det perifera nervsystemet och immunsystemet anses traditionellt tjäna separata funktioner. Denna linje blir dock alltmer suddig av nya insikter om neurogen inflammation. Nociceptorneuroner har många av samma molekylära igenkänningsvägar för fara som immunceller och som svar på fara kommunicerar det perifera nervsystemet direkt med immunsystemet och bildar en integrerad skyddsmekanism. Det täta innervationsnätverket av sensoriska och autonoma fibrer i perifera vävnader och den höga hastigheten på neural transduktion möjliggör snabb lokal och systemisk neurogen modulering av immunitet. Perifera neuroner verkar också spela en betydande roll i immundysfunktion vid autoimmuna och allergiska sjukdomar. Därför kan förståelse av den samordnade interaktionen av perifera neuroner med immunceller främja terapeutiska metoder för att öka värdförsvaret och undertrycka immunopatologi.
Beskrivning
För två tusen år sedan definierade Celsus inflammation som involverande fyra kardinaltecken � Dolor (smärta), Calor (värme), Rubor (rodnad) och tumör (svullnad), en observation som indikerar att aktivering av nervsystemet ansågs vara en integrerad del av nervsystemet. inflammation. Smärta har dock främst ansetts sedan dess, bara som ett symptom, och inte en deltagare i genereringen av inflammation. I detta perspektiv visar vi att det perifera nervsystemet spelar en direkt och aktiv roll för att modulera medfödd och adaptiv immunitet, så att immun- och nervsystemet kan ha en gemensam integrerad skyddsfunktion i värdförsvaret och svaret på vävnadsskada, en intrikat interaktion som också kan leda till patologi vid allergiska och autoimmuna sjukdomar.
Organismers överlevnad är kritiskt beroende av förmågan att montera ett försvar mot potentiell skada från vävnadsskada och infektion. Värdförsvar involverar både undvikande beteende för att ta bort kontakt med en farlig (skadlig) miljö (en neural funktion), och aktiv neutralisering av patogener (en immunfunktion). Traditionellt har immunsystemets roll för att bekämpa infektionsämnen och reparera vävnadsskada ansetts vara ganska skild från nervsystemets, som omvandlar skadliga miljö- och inre signaler till elektrisk aktivitet för att producera förnimmelser och reflexer (Fig. 1). Vi föreslår att dessa två system faktiskt är komponenter i en enhetlig försvarsmekanism. Det somatosensoriska nervsystemet är idealiskt placerat för att upptäcka fara. För det första, alla vävnader som är mycket exponerade för den yttre miljön, såsom epitelytor i huden, lungorna, urinvägarna och matsmältningsorganen, är tätt innerverade av nociceptorer, smärtproducerande sensoriska fibrer med hög tröskel. För det andra är transduktion av skadliga yttre stimuli nästan omedelbar, storleksordningar snabbare än mobiliseringen av det medfödda immunsystemet, och kan därför vara den första som svarar på värdförsvaret.
Figur 1: Skadliga stimuli, mikrobiella och inflammatoriska igenkänningsvägar utlöser aktivering av det perifera nervsystemet. Sensoriska neuroner har flera sätt att upptäcka närvaron av skadliga/skadliga stimuli. 1) Farosignalreceptorer, inklusive TRP-kanaler, P2X-kanaler och DAMP-receptorer (dangerassociated molecular pattern) känner igen exogena signaler från omgivningen (t.ex. värme, surhet, kemikalier) eller endogena farosignaler som frigörs under trauma/vävnadsskada (t.ex. ATP, urinsyra, hydroxynonenaler). 2) Mönsterigenkänningsreceptorer (PRR) såsom Toll-like receptors (TLRs) och Nod-like receptors (NLRs) känner igen patogenassocierade molekylära mönster (PAMPs) som avges genom att invadera bakterier eller virus under infektion. 3) Cytokinreceptorer känner igen faktorer som utsöndras av immunceller (t.ex. IL-1beta, TNF-alfa, NGF), som aktiverar kartkinaser och andra signalmekanismer för att öka membranexcitabilitet.
Förutom ortodroma input till ryggmärgen och hjärnan från periferin kan aktionspotentialer i nociceptorneuroner även överföras antidromiskt vid grenpunkter tillbaka ner till periferin, axonreflexen. Dessa tillsammans med ihållande lokala depolariseringar leder till en snabb och lokal frisättning av neurala mediatorer från både perifera axoner och terminaler (Fig. 2) 1. Klassiska experiment av Goltz (1874) och av Bayliss (1901) visade att elektriskt stimulerande dorsala rötter inducerar hudvasodilatation, vilket ledde till konceptet av en �neurogen inflammation�, oberoende av den som produceras av immunsystemet (Fig. 3).
Figur 2: Neuronala faktorer som frigörs från nociceptorsensoriska neuroner driver direkt leukocytkemotaxi, vaskulär hemodynamik och immunsvaret. När skadliga stimuli aktiverar afferenta signaler i sensoriska nerver, genereras antidromiska axonreflexer som inducerar frisättning av neuropeptider vid neuronernas perifera terminaler. Dessa molekylära mediatorer har flera inflammatoriska effekter: 1) Kemotaxi och aktivering av neutrofiler, makrofager och lymfocyter till platsen för skadan och degranulering av mastceller. 2) Signalering till vaskulära endotelceller för att öka blodflödet, vaskulärt läckage och ödem. Detta möjliggör också enklare rekrytering av inflammatoriska leukocyter. 3) Priming av dendritiska celler för att driva efterföljande T-hjälparcellsdifferentiering till Th2- eller Th17-subtyper.
Figur 3: Tidslinje för framsteg i förståelsen av de neurogena aspekterna av inflammation från Celsus till idag.
Neurogen inflammation medieras av frisättningen av neuropeptiderna kalcitoningenrelaterad peptid (CGRP) och substans P (SP) från nociceptorer, som verkar direkt på vaskulära endotelceller och glatta muskelceller 2�5. CGRP ger vasodilatationseffekter 2, 3, medan SP ökar kapillärpermeabiliteten vilket leder till plasmaextravasation och ödem 4, 5, vilket bidrar till rubor, kalori och tumör i Celsus. Emellertid frisätter nociceptorer många ytterligare neuropeptider (onlinedatabas: www.neuropeptides.nl/), inklusive Adrenomedullin, Neurokinin A och B, Vasoactive intestinal peptide (VIP), neuropeptide (NPY) och gastrin-releasing peptide (GRP), såväl som andra molekylära mediatorer som glutamat, kväveoxid (NO) och cytokiner som eotaxin 6.
Vi inser nu att mediatorerna som frigörs från sensoriska neuroner i periferin inte bara verkar på kärlsystemet, utan också direkt attraherar och aktiverar medfödda immunceller (mastceller, dendritiska celler) och adaptiva immunceller (T-lymfocyter) 7�12. I den akuta situationen med vävnadsskada antar vi att neurogen inflammation är skyddande, underlättar fysiologisk sårläkning och immunförsvar mot patogener genom att aktivera och rekrytera immunceller. Sådana neuroimmuna kommunikationer spelar emellertid sannolikt också viktiga roller i patofysiologin för allergiska och autoimmuna sjukdomar genom att förstärka patologiska eller maladaptiva immunsvar. I djurmodeller av reumatoid artrit till exempel, har Levine och kollegor visat att denervering av leden leder till en slående dämpning av inflammation, vilket är beroende av neuralt uttryck av substans P 13, 14. I nyare studier av allergisk luftvägsinflammation, kolit och psoriasis, primära sensoriska neuroner spelar en central roll i att initiera och förstärka aktiveringen av medfödd och adaptiv immunitet 15�17.
Vi föreslår därför att det perifera nervsystemet inte bara spelar en passiv roll i värdförsvaret (detektering av skadliga stimuli och initiering av undvikandebeteende), utan också en aktiv roll i samverkan med immunsystemet för att modulera svaren på och bekämpa skadliga stimuli, en roll som kan undergrävas för att bidra till sjukdom.
Delade faroidentifieringsvägar i det perifera nervsystemet och det medfödda immunsystemet
Perifera sensoriska neuroner är anpassade för att känna igen fara för organismen på grund av deras känslighet för intensiva mekaniska, termiska och irriterande kemiska stimuli (Fig. 1). Transient receptor potential (TRP) jonkanaler är de mest studerade molekylära mediatorerna av nociception, som utför icke-selektivt inträde av katjoner vid aktivering av olika skadliga stimuli. TRPV1 aktiveras av höga temperaturer, lågt pH och capsaicin, den vallinoidirriterande komponenten i chilipeppar 18. TRPA1 förmedlar detektering av reaktiva kemikalier inklusive miljöirriterande ämnen som tårgas och industriella isotiocyanater 19, men ännu viktigare, det aktiveras också under vävnad skada av endogena molekylära signaler inklusive 4-hydroxinonenal och prostaglandiner 20, 21.
Intressant nog delar sensoriska neuroner många av samma patogen- och faramolekylära igenkänningsreceptorvägar som medfödda immunceller, vilket gör att de också kan upptäcka patogener (Fig. 1). I immunsystemet detekteras mikrobiella patogener av könslinjekodade mönsterigenkänningsreceptorer (PRR), som känner igen brett konserverade exogena patogenassocierade molekylära mönster (PAMP). De första PRR som identifierades var medlemmar av familjen toll-like receptor (TLR), som binder till jäst, bakteriehärledda cellväggskomponenter och viralt RNA 22. Efter PRR-aktivering aktiveras nedströms signalvägar som inducerar cytokinproduktion och aktivering av adaptiv immunitet. Förutom TLR aktiveras medfödda immunceller under vävnadsskada av endogent härledda farosignaler, även kända som skadeassocierade molekylära mönster (DAMPs) eller alarminer 23, 24. Dessa farosignaler inkluderar HMGB1, urinsyra och värmechockproteiner som frigörs genom att dö celler under nekros, aktivera immunceller under icke-infektiösa inflammatoriska svar.
PRR inklusive TLR 3, 4, 7 och 9 uttrycks av nociceptorneuroner, och stimulering av TLR-ligander leder till induktion av inåtriktade strömmar och sensibilisering av nociceptorer för andra smärtstimuli 25�27. Dessutom leder aktivering av sensoriska neuroner av TLR7-liganden imiquimod till aktivering av en klådaspecifik sensorisk väg 25. Dessa resultat indikerar att infektionsrelaterad smärta och klåda delvis kan bero på direkt aktivering av neuroner av patogen-härledda faktorer, som i sin tur aktivera immunceller genom perifer frisättning av neuronala signalmolekyler.
En viktig DAMP/alarmin som frisätts under cellulär skada är ATP, som känns igen av purinerga receptorer på både nociceptorneuroner och immunceller 28�30. Purinerga receptorer är uppbyggda av två familjer: P2X-receptorer, ligandstyrda katjonkanaler och P2Y-receptorer, G-proteinkopplade receptorer. I nociceptorneuroner sker igenkänning av ATP genom P2X3, vilket leder till snabbt densensibiliserande katjonströmmar och smärta 28, 30 (Fig. 1), medan P2Y-receptorer bidrar till nociceptoraktivering genom sensibilisering av TRP och spänningsstyrda natriumkanaler. I makrofager leder ATP-bindning till P2X7-receptorer till hyperpolarisering och nedströms aktivering av inflammasomen, ett molekylärt komplex som är viktigt för generering av IL-1beta och IL-18 29. Därför är ATP en potent farosignal som aktiverar både perifera neuroner och medfödda immunitet under skada, och vissa bevis tyder till och med på att neuroner uttrycker delar av det inflammasommolekylära maskineriet 31.
Baksidan av farosignaler i nociceptorer är rollen av TRP-kanaler i immuncellsaktivering. TRPV2, en homolog av TRPV1 aktiverad av skadlig värme, uttrycks i höga nivåer i medfödda immunceller 32. Genetisk ablation av TRPV2 ledde till defekter i makrofagfagocytos och eliminering av bakterieinfektioner 32. Mastceller uttrycker också TRPV-kanaler, som direkt kan förmedla deras degranulering 33. Det återstår att fastställa om endogena farosignaler aktiverar immunceller på ett liknande sätt som nociceptorer.
Ett nyckelsätt för kommunikation mellan immunceller och nociceptorneuroner är genom cytokiner. Vid aktivering av cytokinreceptorer aktiveras signaltransduktionsvägar i sensoriska neuroner, vilket leder till nedströms fosforylering av membranproteiner inklusive TRP och spänningsstyrda kanaler (Fig. 1). Den resulterande sensibiliseringen av nociceptorer innebär att normalt ofarliga mekaniska stimuli och värmestimuli nu kan aktivera nociceptorer. Interleukin 1 beta och TNF-alfa är två viktiga cytokiner som frisätts av medfödda immunceller under inflammation. IL-1beta och TNF-alfa avkänns direkt av nociceptorer som uttrycker de besläktade receptorerna, inducerar aktivering av p38 map kinaser vilket leder till ökad membranexcitabilitet 34�36. Nervtillväxtfaktor (NGF) och prostaglandin E(2) är också viktiga inflammatoriska mediatorer som frigörs från immunceller som verkar direkt på perifera sensoriska neuroner för att orsaka sensibilisering. En viktig effekt av nociceptorsensibilisering av immunfaktorer är en ökad frisättning av neuropeptider vid perifera terminaler som ytterligare aktiverar immunceller och därigenom inducerar en positiv återkopplingsloop som driver och underlättar inflammation.
Sensoriskt nervsystem kontroll av medfödd och adaptiv immunitet
I tidiga faser av inflammation signalerar sensoriska neuroner till vävnadsboende mastceller och dendritiska celler, som är medfödda immunceller som är viktiga för att initiera immunsvaret (Fig. 2). Anatomiska studier har visat en direkt sammansättning av terminaler med mastceller, såväl som med dendritiska celler, och neuropeptiderna som frigörs från nociceptorer kan inducera degranulering eller cytokinproduktion i dessa celler 7, 9, 37. Denna interaktion spelar en viktig roll i allergiska luftvägar. inflammation och dermatit 10�12.
Under effektorfasen av inflammation måste immuncellerna hitta vägen till den specifika skadan. Många mediatorer som frigörs från sensoriska neuroner, neuropeptider, kemokiner och glutamat, är kemotaktiska för neutrofiler, eosinofiler, makrofager och T-celler och förbättrar endoteladhesion vilket underlättar immuncellernas målsökning 6, 38�41 (Fig. 2). Dessutom tyder vissa bevis på att neuroner kan delta direkt i effektorfasen, eftersom neuropeptider själva kan ha direkta antimikrobiella funktioner 42.
Neuronalt härledda signalmolekyler kan också styra typen av inflammation, genom att bidra till differentiering eller specifikation av olika typer av adaptiva immun-T-celler. Ett antigen fagocyteras och bearbetas av medfödda immunceller, som sedan migrerar till närmaste lymfkörtel och presenterar den antigena peptiden för nava T-celler. Beroende på typen av antigen, samstimulerande molekyler på den medfödda immuncellen och kombinationerna av specifika cytokiner, mognar nava T-celler till specifika subtyper som bäst tjänar den inflammatoriska ansträngningen att rensa den patogena stimulansen. CD4 T-celler, eller T-hjälparceller (Th), kan delas in i fyra huvudgrupper, Th1, Th2, Th17 och T-regulatoriska celler (Treg). Th1-celler är huvudsakligen involverade i att reglera immunsvar mot intracellulära mikroorganismer och organspecifika autoimmuna sjukdomar; Th2 är kritiska för immunitet mot extracellulära patogener, såsom helminter, och är ansvariga för allergiska inflammatoriska sjukdomar; Th17-celler spelar en central roll i skyddet mot mikrobiella utmaningar, såsom extracellulära bakterier och svampar; Treg-celler är involverade i att upprätthålla självtolerans och reglera immunsvar. Denna T-cellmognadsprocess verkar vara starkt påverkad av sensoriska neuronala mediatorer. Neuropeptider, såsom CGRP och VIP, kan påverka dendritiska celler mot en Th2-typ immunitet och minska Th1-typ immunitet genom att främja produktionen av vissa cytokiner och hämma andra, samt genom att minska eller förstärka migration av dendritiska celler till lokala lymfkörtlar 8 , 10, 43. Sensoriska neuroner bidrar också avsevärt till allergisk (främst Th2-driven) inflammation 17. Förutom att reglera Th1- och Th2-celler kan andra neuropeptider, såsom SP och Hemokinin-1, driva det inflammatoriska svaret mer mot Th17 eller Treg 44, 45, vilket betyder att neuroner också kan vara inblandade i att reglera inflammatorisk upplösning. I immunopatologier som kolit och psoriasis kan blockad av neuronala mediatorer som substans P avsevärt dämpa T-cells- och immunförmedlad skada 15�17, även om antagonisering av en mediator i sig bara kan ha en begränsad effekt på neurogen inflammation.
Med tanke på att signalmolekyler som frigörs från perifera sensoriska nervfibrer inte bara reglerar små blodkärl, utan också kemotaxi, målsökning, mognad och aktivering av immunceller, blir det tydligt att neuroimmuna interaktioner är mycket mer komplicerade än man tidigare trott (Fig. . 2). Dessutom är det ganska tänkbart att det inte är individuella neurala mediatorer utan snarare specifika kombinationer av signalmolekyler som frigörs från nociceptorer som påverkar olika stadier och typer av immunsvar.
Autonomisk reflexkontroll av immunitet
En roll för en kolinerg autonoma nervsystemets �reflex�-krets i regleringen av perifera immunsvar förefaller också framträdande 46. Vagusen är den parasympatiska huvudnerven som förbinder hjärnstammen med viscerala organ. Arbete av Kevin Tracey och andra pekar på potenta generaliserade antiinflammatoriska svar vid septisk chock och endotoxemi, utlöst av en efferent vagal nervaktivitet som leder till ett undertryckande av perifera makrofager 47�49. Vagusen aktiverar perifera adrenerga celiaki ganglieneuroner som innerverar mjälten, vilket leder till nedströms frisättning av acetylkolin, som binder till alfa-7 nikotinreceptorer på makrofager i mjälten och mag-tarmkanalen. Detta inducerar aktivering av JAK2/STAT3 SOCS3-signalvägen, som kraftfullt undertrycker TNF-alfa-transkription 47. Det adrenerga celiaki-gangliet kommunicerar också direkt med en undergrupp av acetylkolinproducerande minnes-T-celler, som undertrycker inflammatoriska makrofager 48.
Invarianta naturliga Killer T-celler (iNKT) är en specialiserad undergrupp av T-celler som känner igen mikrobiella lipider i sammanhanget av CD1d istället för peptidantigener. NKT-celler är en nyckel-lymfocytpopulation involverad i kampen mot infektiösa patogener och reglering av systemisk immunitet. NKT-celler finns och trafikerar huvudsakligen genom kärlsystemet och sinusoiderna i mjälten och levern. Sympatiska beta-adrenerga nerver i levern signalerar direkt att modulera NKT-cellaktivitet 50. Under en musmodell av stroke (MCAO), till exempel, undertrycktes lever-NKT-cellernas rörlighet synbart, vilket vändes av sympatisk denervering eller beta-adrenerga antagonister. Dessutom ledde denna immunsuppressiva aktivitet av noradrenerga neuroner på NKT-celler till ökningar av systemisk infektion och lungskada. Därför kan efferenta signaler från autonoma neuroner förmedla en potent immunsuppression.
Dr Alex Jimenez insikt
Neurogen inflammation är en lokal inflammatorisk respons som genereras av nervsystemet. Det tros spela en grundläggande roll i patogenesen av en mängd olika hälsoproblem, inklusive migrän, psoriasis, astma, fibromyalgi, eksem, rosacea, dystoni och multipel kemisk känslighet. Även om neurogen inflammation associerad med det perifera nervsystemet har undersökts omfattande, behöver begreppet neurogen inflammation inom det centrala nervsystemet fortfarande forskning. Enligt flera forskningsstudier tros dock magnesiumbrist vara den främsta orsaken till neurogen inflammation. Följande artikel visar en översikt över mekanismerna för neurogen inflammation i nervsystemet, vilket kan hjälpa vårdpersonal att avgöra det bästa behandlingssättet för att ta hand om en mängd olika hälsoproblem som är förknippade med nervsystemet.
Slutsatser
Vilka är de respektive specifika rollerna för de somatosensoriska och autonoma nervsystemen för att reglera inflammation och immunsystemet (Fig. 4)? Aktivering av nociceptorer leder till lokala axonreflexer, som lokalt rekryterar och aktiverar immunceller och är därför huvudsakligen pro-inflammatoriska och rumsligt begränsade. Däremot leder autonom stimulering till en systemisk immunsuppression genom att påverka pooler av immunceller i lever och mjälte. De afferenta signalmekanismerna i periferin som leder till utlösandet av den immunsuppressiva vagala kolinerga reflexkretsen är dåligt förstådda. Emellertid är 80–90 % av vagala fibrer primära afferenta sensoriska fibrer, och därför kan signaler från inälvorna, många potentiellt drivna av immunceller, leda till aktivering av interneuroner i hjärnstammen och genom dem till en produktion i efferenta vagala fibrer 46.
Figur 4: Sensoriska och autonoma nervsystem modulerar lokala respektive systemiska immunsvar. Nociceptorer som innerverar epitelytor (t.ex. hud och lungor) inducerar lokaliserade inflammatoriska svar, aktiverar mastceller och dendritiska celler. Vid allergisk luftvägsinflammation, dermatit och reumatoid artrit spelar nociceptorneuroner en roll för att driva inflammation. Däremot reglerar autonoma kretsar som innerverar de viscerala organen (t.ex. mjälte och lever) systemiska immunsvar genom att blockera makrofager och NKT-cellaktivering. Vid stroke och septisk endotoxemi spelar dessa neuroner en immunsuppressiv roll.
Typiskt definieras tidsförloppet och arten av inflammation, oavsett om det är under infektion, allergiska reaktioner eller autoimmuna patologier, av kategorierna av immunceller som är involverade. Det kommer att vara viktigt att veta vilka olika typer av immunceller som regleras av sensoriska och autonoma signaler. En systematisk bedömning av vilka mediatorer som kan frigöras från nociceptorer och autonoma neuroner och uttrycket av receptorer för dessa av olika medfödda och adaptiva immunceller kan hjälpa till att lösa denna fråga.
Under evolutionen har liknande molekylära vägar för detektion av faror utvecklats för både medfödd immunitet och nociception även om cellerna har helt olika utvecklingslinjer. Medan PRR och skadliga ligand-styrda jonkanaler studeras separat av immunologer och neurobiologer, är gränsen mellan dessa två områden alltmer suddig. Under vävnadsskada och patogen infektion kommer frisättning av farosignaler sannolikt att leda till en koordinerad aktivering av både perifera neuroner och immunceller med komplex dubbelriktad kommunikation och ett integrerat värdförsvar. Den anatomiska placeringen av nociceptorer i gränssnittet med omgivningen, hastigheten på neurala transduktion och deras förmåga att frigöra potenta cocktails av immunverkande mediatorer tillåter det perifera nervsystemet att aktivt modulera det medfödda immunsvaret och koordinera nedströms adaptiv immunitet. Omvänt är nociceptorer mycket känsliga för immunmediatorer, som aktiverar och sensibiliserar neuronerna. Neurogen och immunförmedlad inflammation är därför inte oberoende enheter utan fungerar tillsammans som anordningar för tidig varning. Men det perifera nervsystemet spelar också en viktig roll i patofysiologin, och kanske etiologin, för många immunsjukdomar som astma, psoriasis eller kolit eftersom dess förmåga att aktivera immunsystemet kan förstärka patologisk inflammation 15�17. Behandling för immunsjukdomar kan därför behöva inkludera målinriktning av nociceptorer såväl som immunceller.
Tack
Vi tackar NIH för stödet (2R37NS039518).
SammanfattningsvisAtt förstå rollen av neurogen inflammation när det kommer till värdförsvar och immunopatologi är avgörande för att bestämma rätt behandlingsmetod för en mängd olika hälsoproblem i nervsystemet. Genom att titta på interaktionerna mellan de perifera neuronerna med immunceller, kan sjukvårdspersonal utveckla terapeutiska metoder för att ytterligare bidra till att öka värdens försvar samt undertrycka immunopatologi. Syftet med artikeln ovan är att hjälpa patienter att förstå neuropatiens kliniska neurofysiologi, bland annat hälsoproblem med nervskador. Information som refereras från National Center for Biotechnology Information (NCBI). Omfattningen av vår information är begränsad till kiropraktik såväl som till ryggradsskador och tillstånd. För att diskutera ämnet är du välkommen att fråga Dr Jimenez eller kontakta oss på�915-850-0900 .
Curated av Dr. Alex Jimenez
Ytterligare ämnen: Ryggsmärta
Ryggont är en av de vanligaste orsakerna till funktionshinder och missade dagar på jobbet över hela världen. Faktum är att ryggsmärta har tillskrivits som den näst vanligaste anledningen till doktorsbesök, som endast överträffas av övre luftvägsinfektioner. Cirka 80 procent av befolkningen kommer att uppleva någon typ av ryggsmärta minst en gång i livet. Ryggraden är en komplex struktur bestående av ben, leder, ligament och muskler, bland annat mjuka vävnader. På grund av detta, skador och / eller förvärrade förhållanden, till exempel hernierade skivor, kan så småningom leda till symtom på ryggont. Sportskador eller skador på bilolyckor är ofta den vanligaste orsaken till ryggsmärta, men ibland kan de enklaste rörelserna få ont. Lyckligtvis kan alternativa behandlingsalternativ, som kiropraktikvård, hjälpa till att lindra ryggont genom användningen av ryggmärgsjusteringar och manuell manipulation, vilket i slutändan förbättrar smärtlindringen.
1.�Sauer SK, Reeh PW, Bove GM. Skadlig värmeinducerad CGRP-frisättning från ischiasnervens axoner från råtta in vitro.�Eur J Neurosci.�2001;14:1203�1208.�[PubMed]
2.�Edvinsson L, Ekman R, Jansen I, McCulloch J, Uddman R. Calcitonin genrelaterad peptid och cerebrala blodkärl: distribution och vasomotoriska effekter.�J Cereb Blood Flow Metab.�1987;7:720�728.�[PubMed]
3.�McCormack DG, Mak JC, Coupe MO, Barnes PJ. Kalcitoningenrelaterad peptidvasodilatation av mänskliga lungkärl.�J Appl Physiol.�1989;67:1265�1270.�[PubMed]
4.�Saria A. Substans P i sensoriska nervfibrer bidrar till utvecklingen av ödem i råttans baktass efter termisk skada.�Br J Pharmacol.�1984;82:217�222.�[PMC gratis artikel]�[PubMed]
5.�Brain SD, Williams TJ. Interaktioner mellan takykininerna och den kalcitoningenererade peptiden leder till modulering av ödembildning och blodflöde i råtthud.�Br J Pharmacol.�1989;97:77�82.[PMC gratis artikel]�[PubMed]
6.�Fryer AD, et al. Neuronalt eotaxin och effekterna av CCR3-antagonist på luftvägshyperreaktivitet och M2-receptordysfunktion.�J Clin Invest.�2006;116:228�236.�[PMC gratis artikel]�[PubMed]
7.�Ansel JC, Brown JR, Payan DG, Brown MA. Substans P aktiverar selektivt TNF-alfa-genuttryck i murina mastceller.�J Immunol.�1993;150:4478�4485.�[PubMed]
9.�Hosoi J, et al. Reglering av Langerhans cellfunktion av nerver som innehåller kalcitoningenrelaterad peptid.�Naturen.�1993;363:159�163.�[PubMed]
10.�Mikami N, et al. Kalcitoningenrelaterad peptid är en viktig regulator av kutan immunitet: effekt på dendritiska celler och T-cellsfunktioner.�J Immunol.�2011;186:6886�6893.�[PubMed]
11.�Rochlitzer S, et al. Den neuropeptid-kalcitoningen-relaterade peptiden påverkar allergisk luftvägsinflammation genom att modulera dendritiska cellfunktionen.�Clin Exp Allergi.�2011;41:1609�1621.�[PubMed]
12.�Cyphert JM, et al. Samarbete mellan mastceller och neuroner är avgörande för antigenmedierad bronkkonstriktion.�J Immunol.�2009;182:7430�7439.�[PMC gratis artikel]�[PubMed]
13.�Levine JD, et al. Intraneuronal substans P bidrar till svårighetsgraden av experimentell artrit.�Vetenskap.�1984;226:547�549.�[PubMed]
14.�Levine JD, Khasar SG, Green PG. Neurogen inflammation och artrit.�Ann NY Acad Sci.�2006;1069:155�167.�[PubMed]
15.�Engel MA, et al. TRPA1 och substans P förmedlar kolit hos möss.�Gastroenterologi.�2011;141:1346�1358.�[PubMed]
16.�Ostrowski SM, Belkadi A, Loyd CM, Diaconu D, Ward NL. Kutan denervering av psoriasismushud förbättrar akantos och inflammation på ett sensoriskt neuropeptidberoende sätt.�J Invest Dermatol.�2011;131:1530�1538.�[PMC gratis artikel]�[PubMed]
17.�Caceres AI, et al. En sensorisk neuronal jonkanal som är avgörande för luftvägsinflammation och hyperreaktivitet vid astma.�Proc Natl Acad Sci US A.�2009;106:9099�9104.�[PMC gratis artikel]�[PubMed]
18.�Caterina MJ, et al. Nedsatt nociception och smärtkänsla hos möss som saknar capsaicinreceptorn.�Vetenskap.�2000;288:306�313.�[PubMed]
19.�Bessac BF, et al. Transient receptorpotential ankyrin 1-antagonister blockerar de skadliga effekterna av giftiga industriella isocyanater och tårgaser.�FASEB J.�2009;23:1102�1114.�[PMC gratis artikel]�[PubMed]
20.�Cruz-Orengo L, et al. Kutan nociception framkallad av 15-delta PGJ2 via aktivering av jonkanalen TRPA1.�Mol smärta.�2008;4:30.�[PMC gratis artikel]�[PubMed]
21.�Trevisani M, et al. 4-Hydroxynonenal, en endogen aldehyd, orsakar smärta och neurogen inflammation genom aktivering av den irriterande receptorn TRPA1.�Proc Natl Acad Sci US A.�2007;104:13519�13524.�[PMC gratis artikel]�[PubMed]
22.�Janeway CA, Jr, Medzhitov R. Inledning: den medfödda immunitetens roll i det adaptiva immunsvaret.�Semin Immunol.�1998;10:349�350.�[PubMed]
23.�Matzinger P. En medfödd känsla av fara.�Ann NY Acad Sci.�2002;961:341�342.�[PubMed]
24.�Bianchi MIG. DAMPs, PAMPs och alarmins: allt vi behöver veta om fara.�J Leukoc Biol.�2007;81:1�5.�[PubMed]
25.�Liu T, Xu ZZ, Park CK, Berta T, Ji RR. Tollliknande receptor 7 förmedlar klåda.�Nat Neurosci.�2010;13:1460�1462.�[PMC gratis artikel]�[PubMed]
26.�Diogenes A, Ferraz CC, Akopian AN, Henry MA, Hargreaves KM. LPS sensibiliserar TRPV1 via aktivering av TLR4 i trigeminussensoriska neuroner.�J Dent Res.�2011;90:759�764.�[PubMed]
27.�Qi J, et al. Smärtsamma vägar inducerade av TLR-stimulering av dorsalrotganglieneuroner.�J Immunol.�2011;186:6417�6426.�[PMC gratis artikel]�[PubMed]
28.�Cockayne DA, et al. Urinblåshyporeflexi och minskat smärtrelaterat beteende hos möss med P2X3-brist.�Naturen.�2000;407:1011�1015.�[PubMed]
29.�Mariathasan S, et al. Kryopyrin aktiverar inflammasomen som svar på toxiner och ATP.�Naturen.�2006;440:228�232.�[PubMed]
30.�Souslova V, et al. Varmkodande brister och avvikande inflammatorisk smärta hos möss som saknar P2X3-receptorer.�Naturen.�2000;407:1015�1017.�[PubMed]
31.�de Rivero Vaccari JP, Lotocki G, Marcillo AE, Dietrich WD, Keane RW. En molekylär plattform i neuroner reglerar inflammation efter ryggmärgsskada.�J Neurosci.�2008;28:3404�3414.�[PubMed]
32.�Link TM, et al. TRPV2 har en avgörande roll i bindning av makrofagpartiklar och fagocytos.�Nat Immunol.�2010;11:232�239.�[PMC gratis artikel]�[PubMed]
33.�Turner H, del Carmen KA, Stokes A. Länk mellan TRPV-kanaler och mastcellsfunktion.�Handb Exp Pharmacol.�2007:457�471.�[PubMed]
34.�Binshtok AM, et al. Nociceptorer är interleukin-1beta-sensorer.�J Neurosci.�2008;28:14062�14073.[PMC gratis artikel]�[PubMed]
35.�Zhang XC, Kainz V, Burstein R, Levy D. Tumörnekrosfaktor-alfa inducerar sensibilisering av meningeala nociceptorer som medieras via lokala COX- och p38 MAP-kinasverkan.�Smärta.�2011;152:140�149.[PMC gratis artikel]�[PubMed]
36.�Samad TA, et al. Interleukin-1beta-medierad induktion av Cox-2 i CNS bidrar till inflammatorisk smärtöverkänslighet.�Naturen.�2001;410:471�475.�[PubMed]
37.�Veres TZ, et al. Rumslig interaktion mellan dendritiska celler och sensoriska nerver vid allergisk luftvägsinflammation.�Am J Respir Cell Mol Biol.�2007;37:553�561.�[PubMed]
38.�Smith CH, Barker JN, Morris RW, MacDonald DM, Lee TH. Neuropeptider inducerar snabbt uttryck av endotelcellsadhesionsmolekyler och framkallar granulocytisk infiltration i mänsklig hud.�J Immunol.�1993;151:3274�3282.�[PubMed]
39.�Dunzendorfer S, Meierhofer C, Wiedermann CJ. Signalering i neuropeptidinducerad migration av humana eosinofiler.�J Leukoc Biol.�1998;64:828�834.�[PubMed]
40.�Ganor Y, Besser M, Ben-Zakay N, Unger T, Levite M. Humana T-celler uttrycker en funktionell jonotropisk glutamatreceptor GluR3, och glutamat triggar i sig integrinmedierad adhesion till laminin och fibronektin och kemotaktisk migration.�J Immunol.�2003;170:4362�4372.�[PubMed]
41.�Czepielewski RS, et al. Gastrin-frisättande peptidreceptor (GRPR) förmedlar kemotaxi i neutrofiler.�Proc Natl Acad Sci US A.�2011;109:547�552.�[PMC gratis artikel]�[PubMed]
42.�Brogden KA, Guthmiller JM, Salzet M, Zasloff M. Nervsystemet och medfödd immunitet: neuropeptidkopplingen.�Nat Immunol.�2005;6:558�564.�[PubMed]
43.�Jimeno R, et al. Effekt av VIP på balansen mellan cytokiner och masterregulatorer av aktiverade T-hjälparceller.�Immunol Cell Biol.�2011;90:178�186.�[PubMed]
44.�Razavi R, et al. TRPV1+ sensoriska neuroner kontrollerar betacellsstress och öinflammation vid autoimmun diabetes.�Cell.�2006;127:1123�1135.�[PubMed]
45.�Cunin P, et al. Tachykininsubstansen P och hemokinin-1 gynnar genereringen av mänskliga minne Th17-celler genom att inducera IL-1beta, IL-23 och TNF-liknande 1A-uttryck av monocyter.�J Immunol.�2011;186:4175�4182.�[PubMed]
46.�Andersson U, Tracey KJ. Reflexprinciper för immunologisk homeostas.�Annu Rev Immunol.�2011[PMC gratis artikel]�[PubMed]
47.�de Jonge WJ, et al. Stimulering av vagusnerven dämpar makrofagaktivering genom att aktivera Jak2-STAT3-signalvägen.�Nat Immunol.�2005;6:844�851.�[PubMed]
48.�Rosas-Ballina M, et al. Acetylkolin-syntetiserande T-celler vidarebefordrar neurala signaler i en vagusnervkrets.�Vetenskap.�2011;334:98�101.�[PMC gratis artikel]�[PubMed]
49.�Wang H, et al. Nikotinacetylkolinreceptorn alfa7 subenhet är en viktig regulator av inflammation.�Naturen.�2003;421:384�388.�[PubMed]
50.�Wong CH, Jenne CN, Lee WY, Leger C, Kubes P. Funktionell innervering av hepatiska iNKT-celler är immunsuppressiv efter stroke.�Vetenskap.�2011;334:101�105.�[PubMed]
IFMs verktyg för Find A Practitioner är det största remissnätverket inom Functional Medicine, skapat för att hjälpa patienter att hitta Functional Medicine-utövare var som helst i världen. IFM Certified Practitioners listas först i sökresultaten, med tanke på deras omfattande utbildning i funktionell medicin



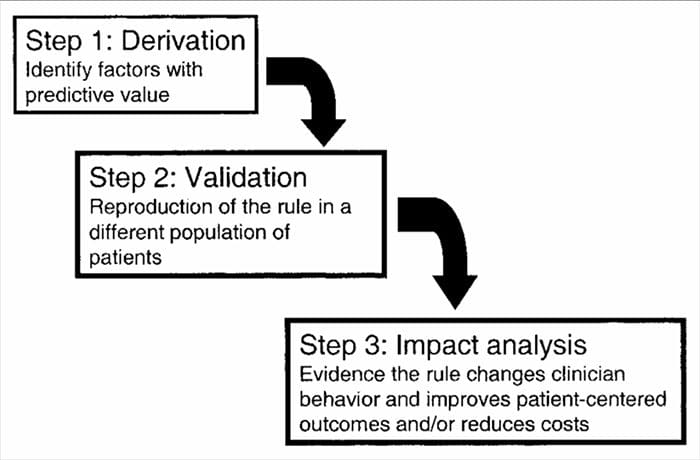




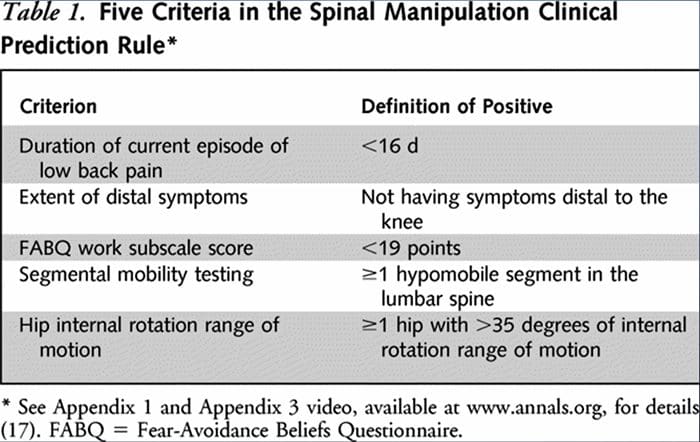
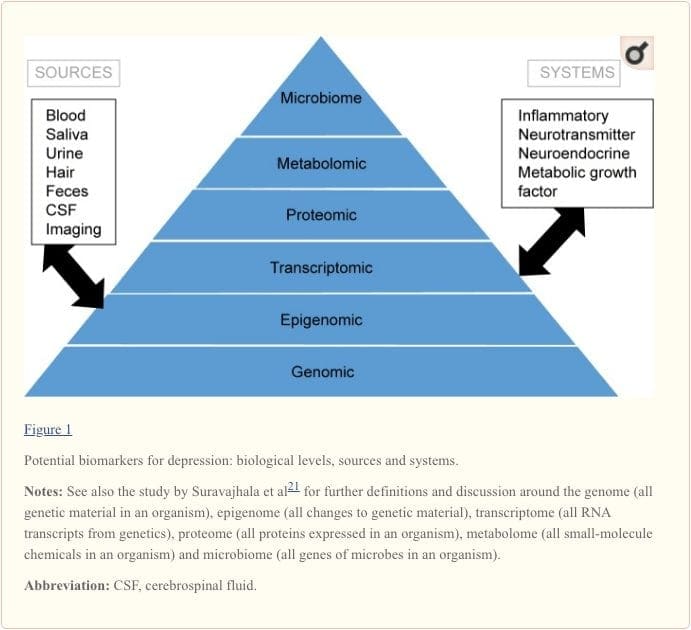





 Abstrakt
Abstrakt

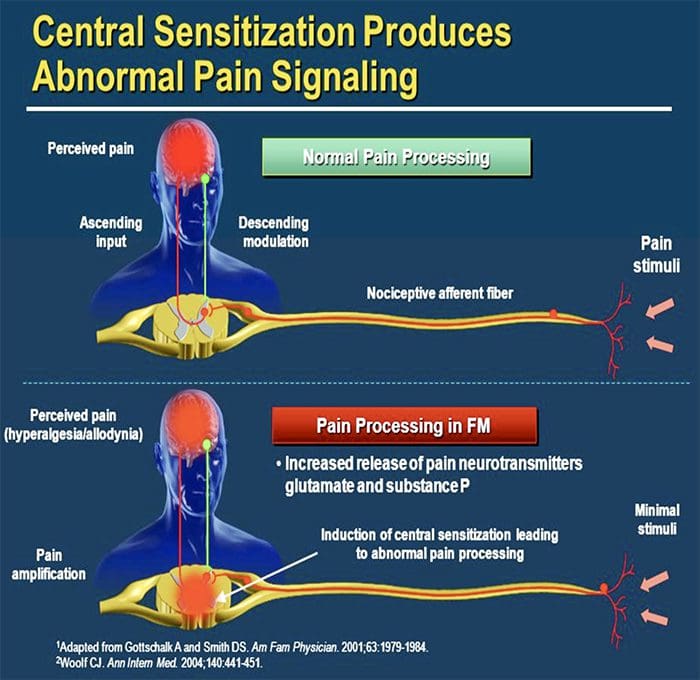















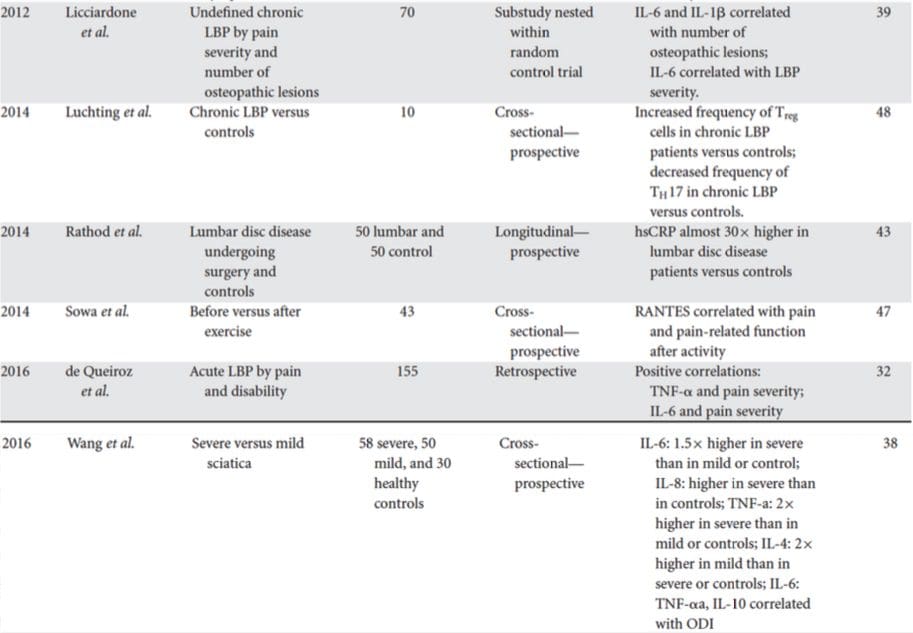


 Referenser:
Referenser:
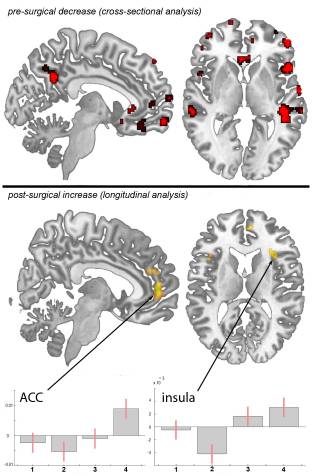
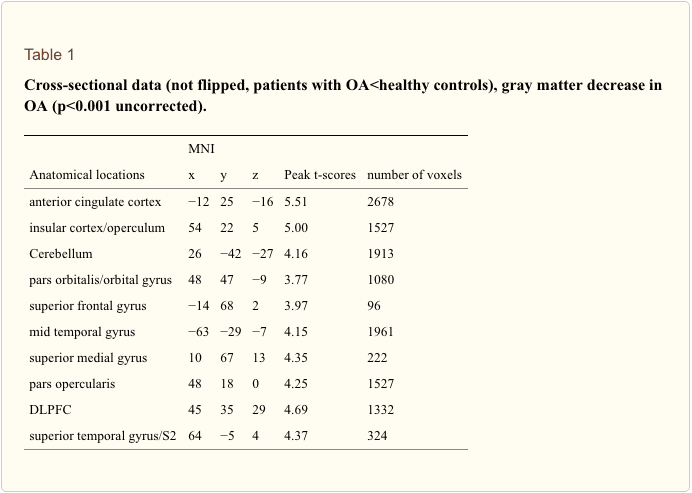
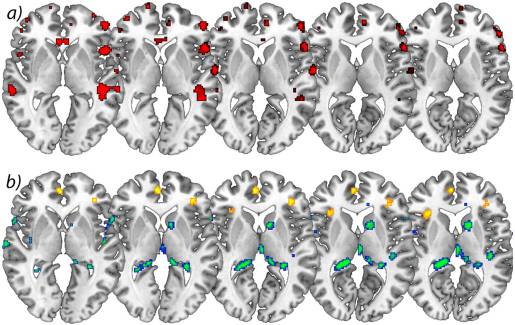
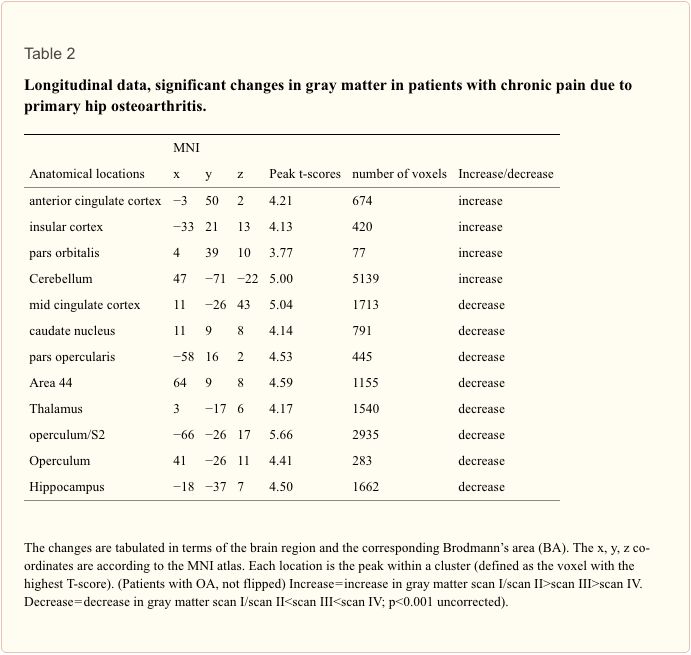
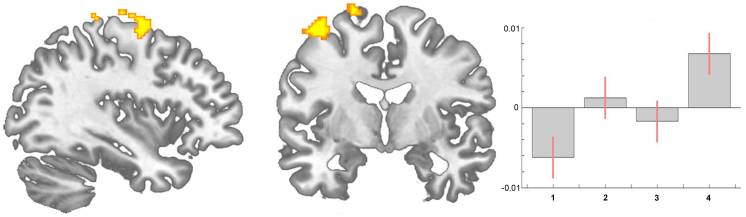
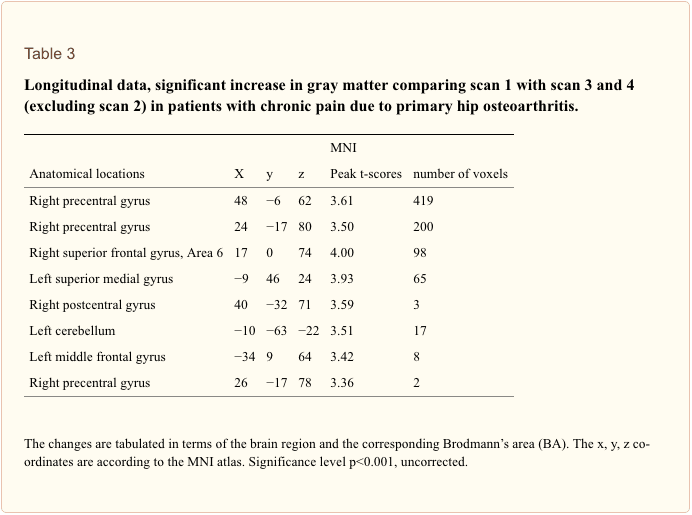

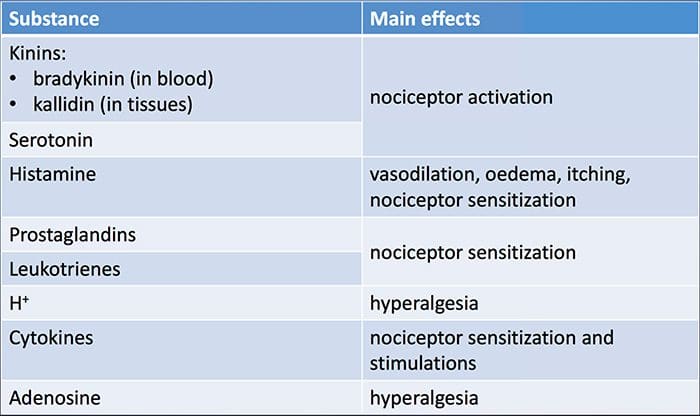
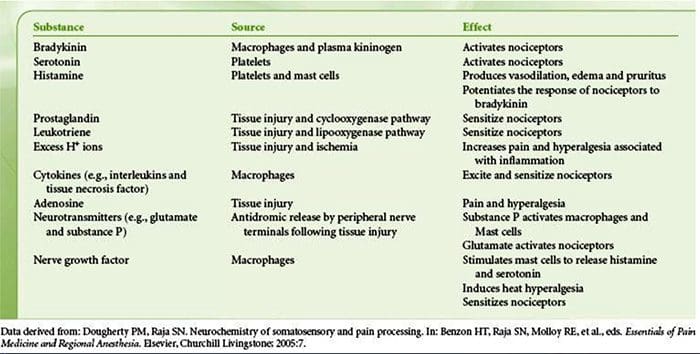
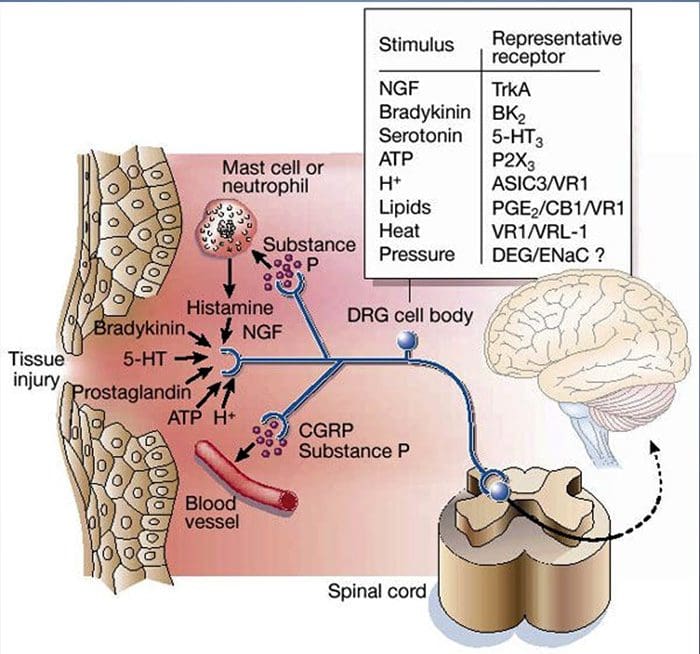
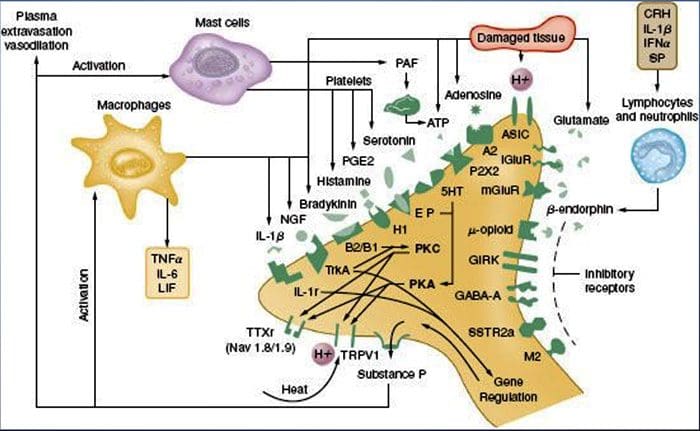 Varför gör min axel ont? En genomgång av den neuroanatomiska och biokemiska grunden för axelsmärta
Varför gör min axel ont? En genomgång av den neuroanatomiska och biokemiska grunden för axelsmärta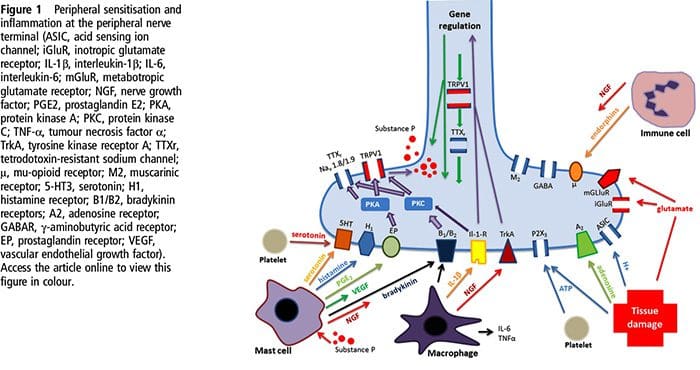 NGF och den transienta receptorpotentiala katjonkanalunderfamiljen V medlem 1 (TRPV1)-receptorn har ett symbiotiskt samband när det kommer till inflammation och nociceptorsensibilisering. Cytokinerna som produceras i inflammerad vävnad resulterar i en ökning av NGF-produktionen.19 NGF stimulerar frisättningen av histamin och serotonin (5-HT3) av mastceller, och sensibiliserar även nociceptorer, vilket möjligen förändrar egenskaperna hos A? fibrer så att en större andel blir nociceptiva. TRPV1-receptorn finns i en subpopulation av primära afferenta fibrer och aktiveras av capsaicin, värme och protoner. TRPV1-receptorn syntetiseras i den afferenta fiberns cellkropp och transporteras till både de perifera och centrala terminalerna, där den bidrar till känsligheten hos nociceptiva afferenter. Inflammation resulterar i NGF-produktion perifert som sedan binder till tyrosinkinasreceptor typ 1-receptorn på nociceptorterminalerna, NGF transporteras sedan till cellkroppen där det leder till en uppreglering av TRPV1-transkription och följaktligen ökad nociceptorkänslighet.19 20 NGF och andra inflammatoriska mediatorer sensibiliserar också TRPV1 genom en mångfald av sekundära budbärarvägar. Många andra receptorer inklusive kolinerga receptorer, a-aminosmörsyra (GABA)-receptorer och somatostatinreceptorer tros också vara involverade i perifer nociceptorkänslighet.
NGF och den transienta receptorpotentiala katjonkanalunderfamiljen V medlem 1 (TRPV1)-receptorn har ett symbiotiskt samband när det kommer till inflammation och nociceptorsensibilisering. Cytokinerna som produceras i inflammerad vävnad resulterar i en ökning av NGF-produktionen.19 NGF stimulerar frisättningen av histamin och serotonin (5-HT3) av mastceller, och sensibiliserar även nociceptorer, vilket möjligen förändrar egenskaperna hos A? fibrer så att en större andel blir nociceptiva. TRPV1-receptorn finns i en subpopulation av primära afferenta fibrer och aktiveras av capsaicin, värme och protoner. TRPV1-receptorn syntetiseras i den afferenta fiberns cellkropp och transporteras till både de perifera och centrala terminalerna, där den bidrar till känsligheten hos nociceptiva afferenter. Inflammation resulterar i NGF-produktion perifert som sedan binder till tyrosinkinasreceptor typ 1-receptorn på nociceptorterminalerna, NGF transporteras sedan till cellkroppen där det leder till en uppreglering av TRPV1-transkription och följaktligen ökad nociceptorkänslighet.19 20 NGF och andra inflammatoriska mediatorer sensibiliserar också TRPV1 genom en mångfald av sekundära budbärarvägar. Många andra receptorer inklusive kolinerga receptorer, a-aminosmörsyra (GABA)-receptorer och somatostatinreceptorer tros också vara involverade i perifer nociceptorkänslighet.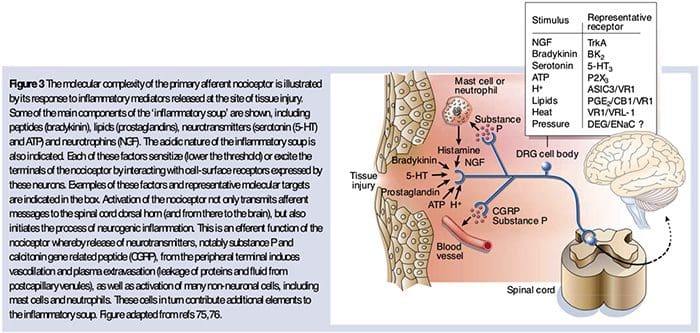 Neurokemin hos nociceptorer
Neurokemin hos nociceptorer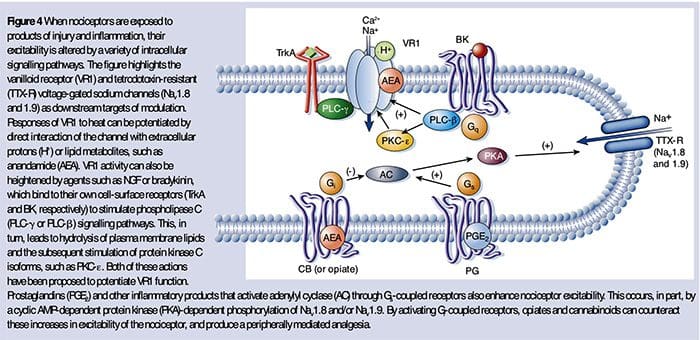 Cellulära och molekylära smärtmekanismer
Cellulära och molekylära smärtmekanismer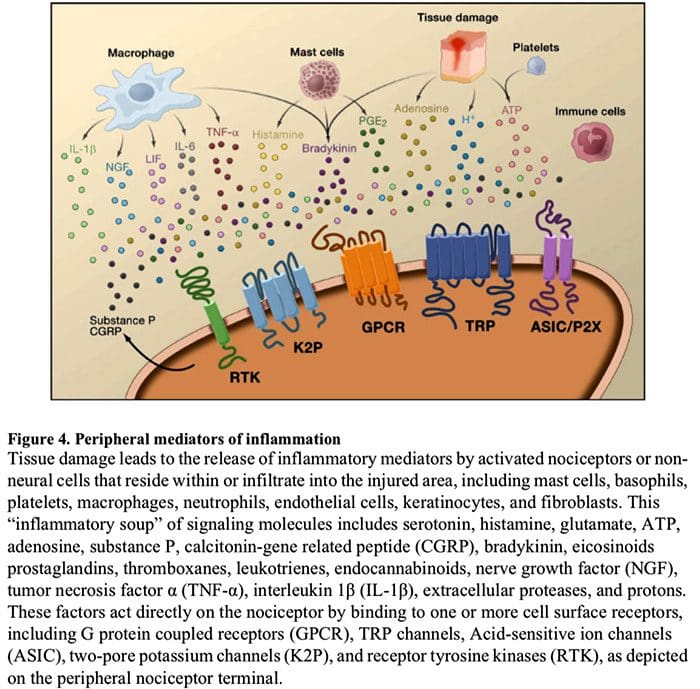
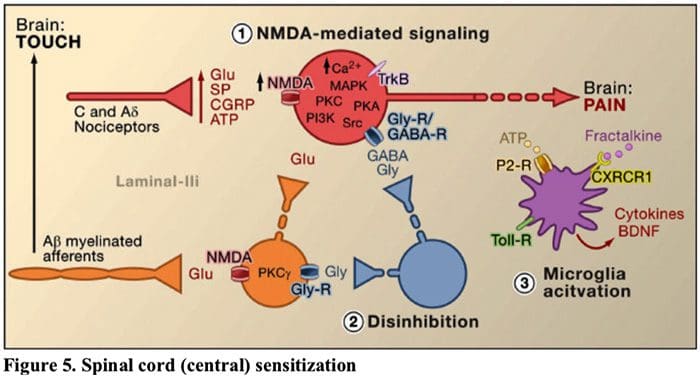 Figur 5. Sensibilisering av ryggmärg (central).
Figur 5. Sensibilisering av ryggmärg (central).